5 Lab Week 5: Basal Ganglia and Cerebellum
Learning Objectives
1. Basal Ganglia
Identify on gross brain specimens, MRIs, and in appropriate sections, the components of the basal ganglia: caudate nucleus, putamen, globus pallidus, substantia nigra, and subthalamic nucleus.
Define the terms: striatum and lentiform nuclei.
Outline the basic looped circuit that links the cortex with the basal ganglia.
Distinguish between the direct and indirect pathways that modulate voluntary motor activity.
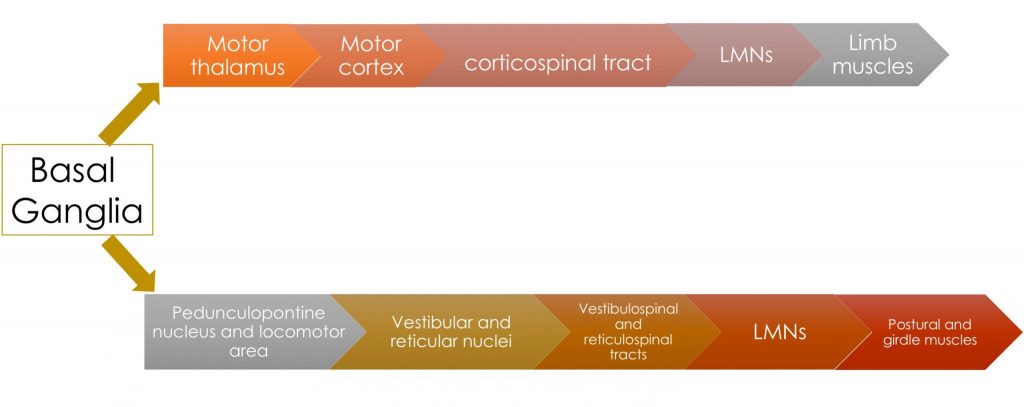
Sketch a flow diagram of the descending pathway from the basal ganglia to the pedunculopontine nucleus in the brainstem, which in turn projects to the nuclei that give rise to the reticulospinal and vestibulospinal tracts.
State which major structures are most affected in the following conditions: hemiballismus, Parkinson disease, Huntington disease.
2. Cerebellum
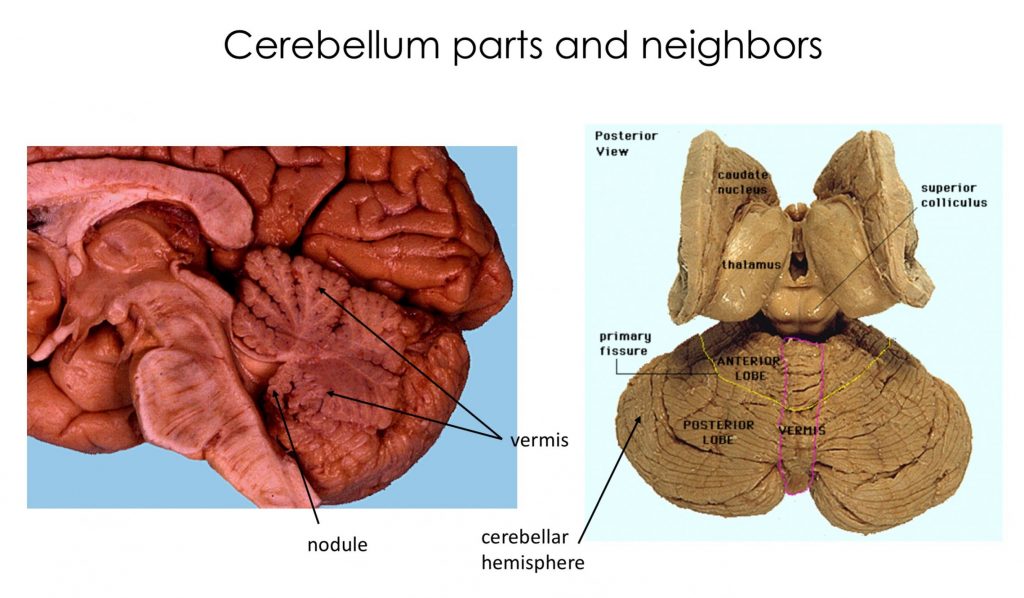
Identify on gross specimens, MRIs and in appropriate sections and dissections: the vermis, tonsil, flocculus, cerebellar hemispheres, dentate nucleus (the only deep cerebellar nucleus easily visible), anterior lobe, posterior lobe, and the cerebellar peduncles.
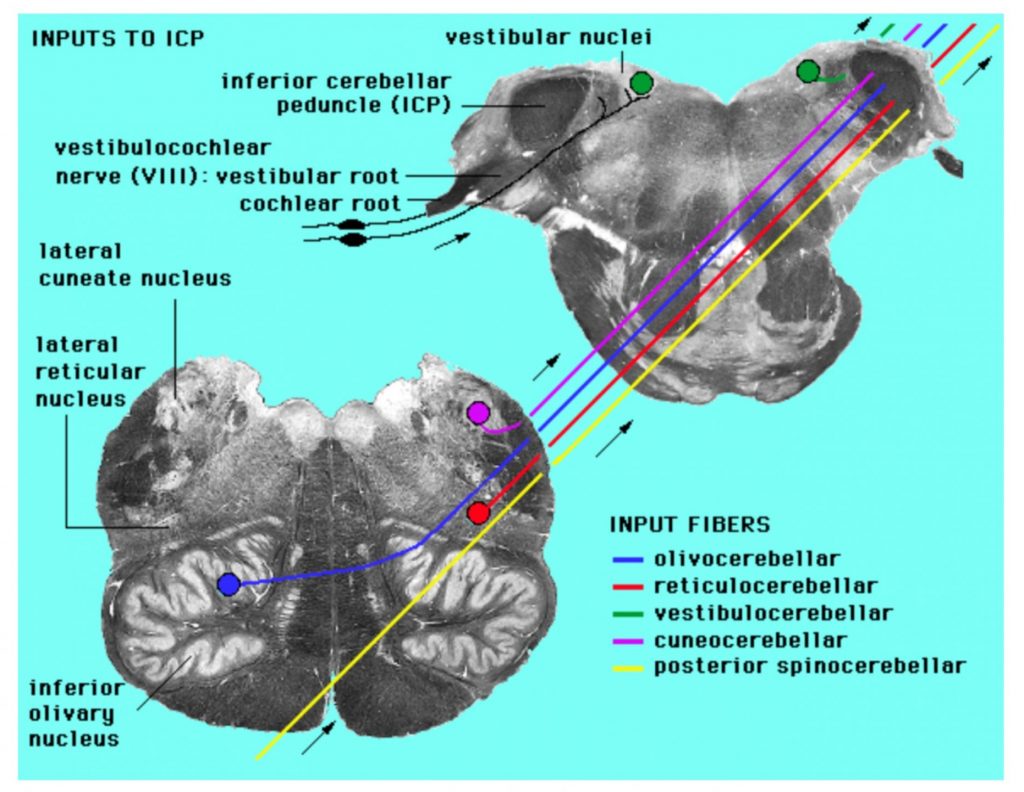
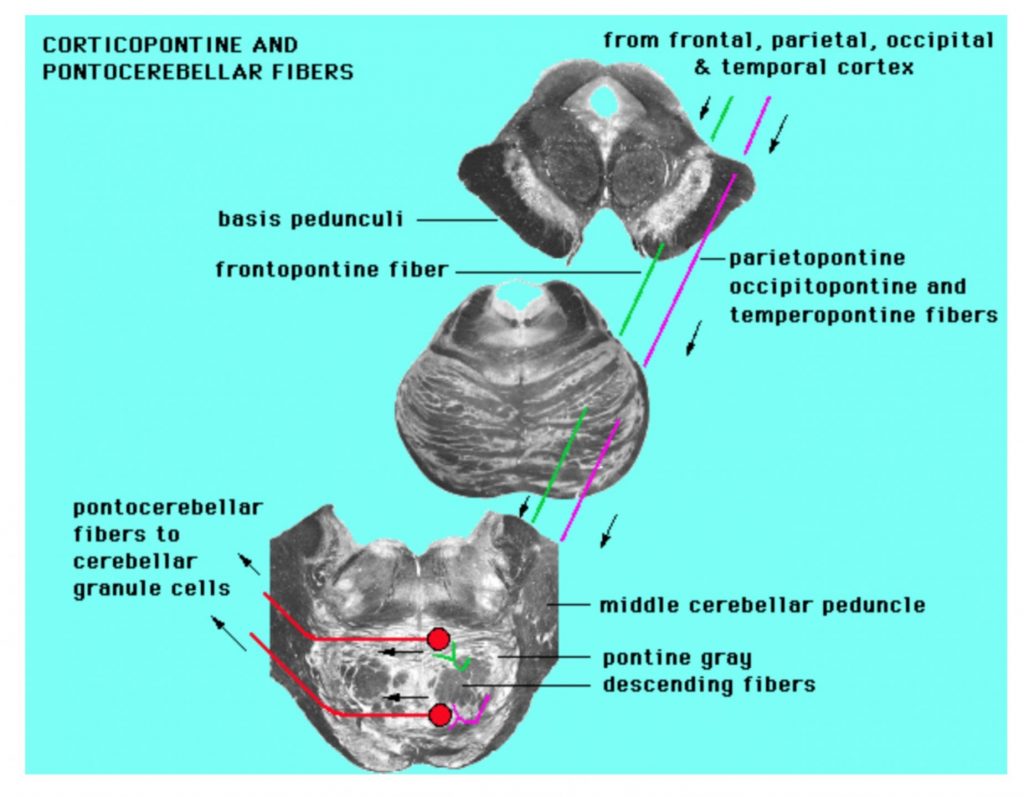
List the inputs to the cerebellum from the spinal cord (dorsal spinocerebellar and cuneocerebellar tracts), brainstem (olivocerebellar, vestibulocerebellar tracts), and cortex via pontine nuclei in terms of origin, presence of crossed fibers, and which cerebellar peduncle they traverse.
Describe the outputs of the cerebellum in terms of brainstem targets (reticular, vestibular and red nuclei) and (eventual) cortex target (i.e. via thalamus).
List some results of damage to the cerebellum.
Overview
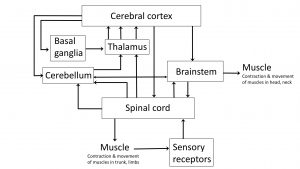
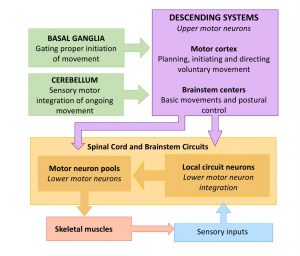
Up to this point, we have focused on structures and circuits that enable voluntary movement and which, when damaged, result in some degree of weakness even to the level of paralysis if the damage is severe. We turn our focus now to other structures in the motor system that enhance voluntary movements, making them more smooth, coordinated, efficient. When these structures are damaged, there is a degradation in the quality of movement, rather than weakness.
In the diagrams below, trace the loops of connection involving the cerebellum and basal ganglia. Notice that while we think of these as being “motor” structures, neither the cerebellum nor basal ganglia have direct connections to motor neurons. Their influence on motor function is mediated through their effect on the cortex (and thus the corticospinal/corticobulbar tracts) and on the reticular and vestibular nuclei (and thus the reticulospinal and vestibulospinal tracts respectively).
Basal Ganglia
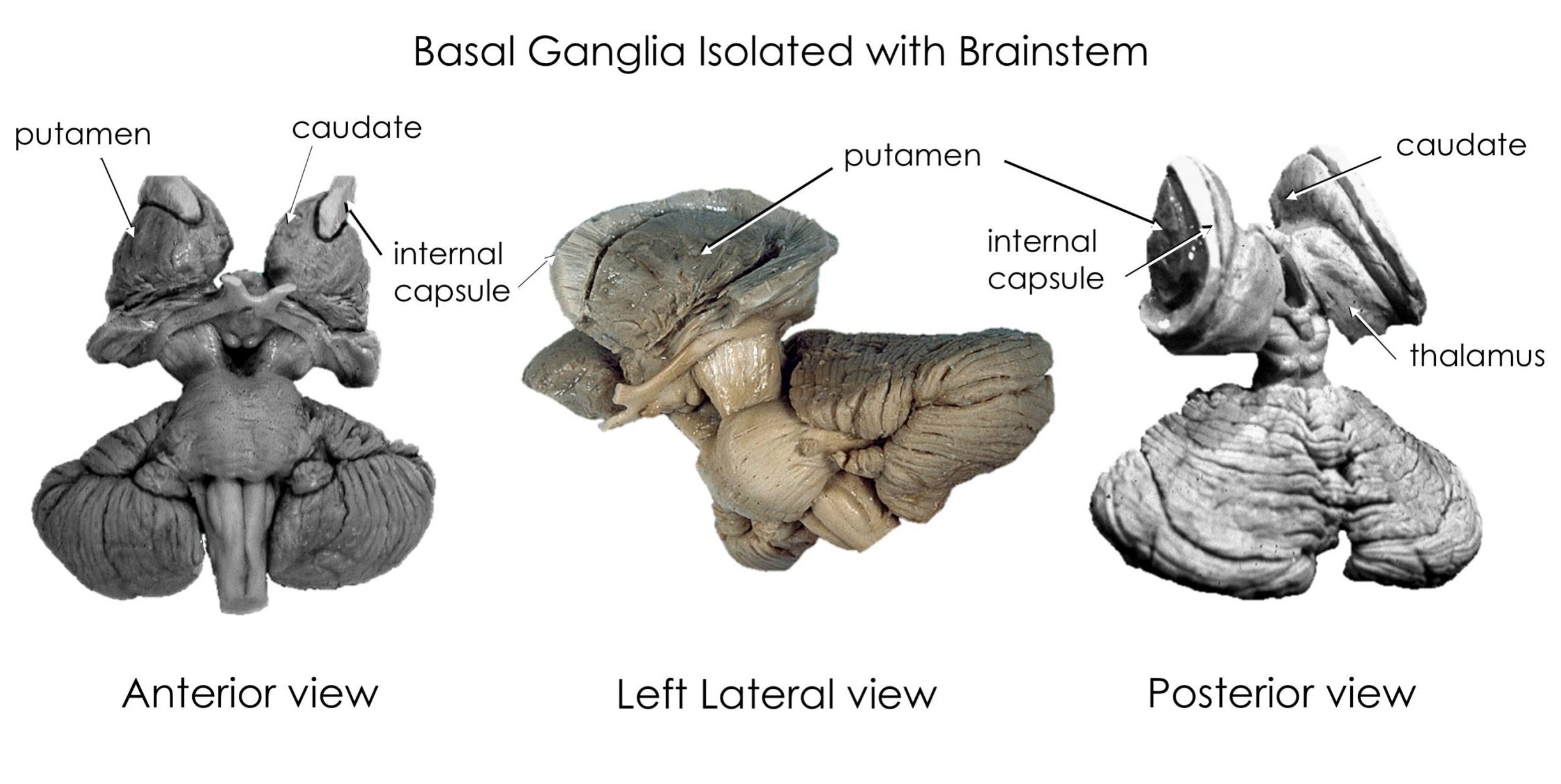
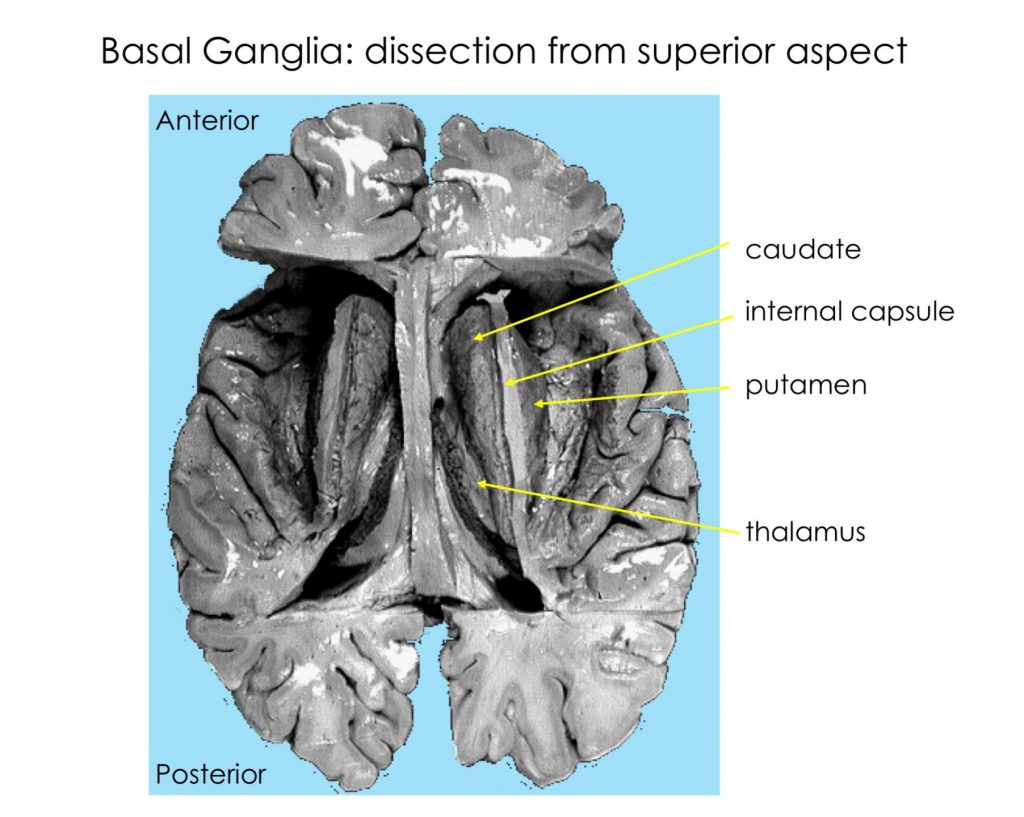
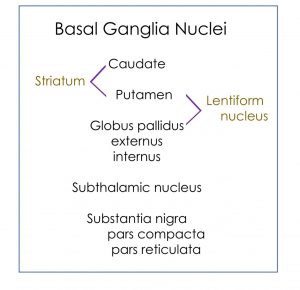 The basal ganglia include structures in the core of the cerebral hemispheres, the diencephalon, and the midbrain. The caudate and putamen are often referred to as the striatum because of their common embryological derivation and their role as input nuclei for the basal ganglia. The globus pallidus and putamen are referred to as the lentiform nucleus in the context of anatomical location.
The basal ganglia include structures in the core of the cerebral hemispheres, the diencephalon, and the midbrain. The caudate and putamen are often referred to as the striatum because of their common embryological derivation and their role as input nuclei for the basal ganglia. The globus pallidus and putamen are referred to as the lentiform nucleus in the context of anatomical location.
The substantia nigra is in the midbrain and the subthalamic nucleus is a nucleus in the diencephalon.
Here are 3 movies that show 3D visualizations of the caudate, putamen and globus pallidus, reconstructed from MRI scans. You might also like to review the more detailed narrated videos in Chapter 2 by Dr. John Sundsten.
This one shows the caudate and putamen. Notice the gap between them? This is where the fibers of the internal capsule pass to and from the cortex and thalamus. The yellow area appearing to connect the caudate and putamen at their most anterior-ventral point is the nucleus accumbens, a nucleus important in the reward pathway and in substance use disorders, but beyond the scope of this course.
This video shows the striatum in relationship to the lateral ventricle. Recall that the 3rd ventricle is surrounded by the thalamus and hypothalamus which both are medial to the basal ganglia.
This video shows a peeling away of the right frontal lobe to demonstrate how the globus pallidus (gray) nestles “inside” the putamen, and is smaller than it. The yellow fiber bundle is the anterior commissure.
Substantia nigra and subthalamic nucleus.
The pars reticulata is an output nucleus of the basal ganglia, projecting to the reticular formation, including the midbrain locomotor region (which contains the pedunculopontine nucleus, cuneiform and subcuneiform nuclei), located in the caudal midbrain tegmentum. Through these projections the SNr can influence posture and gait through the intermediary reticulospinal and vestibulospinal tracts.
Basal ganglia cortical loops and pathways
The basal ganglia are involved in multiple parallel circuit loops involving subsets of cells in cortex, basal ganglia, and thalamus. These circuits regulate many aspects of behavior including voluntary movement, eye movement, motivation, emotional and cognitive behavior. The loops all follow the basic pattern of connection that is
cortex ⇒basal ganglia⇒thalamus⇒cortex
The image below shows 3 of these loops. It is for your interest only. You need to know that such circuits exist, but you need only know the details for the circuit involved in voluntary motor activity as described in the next section.
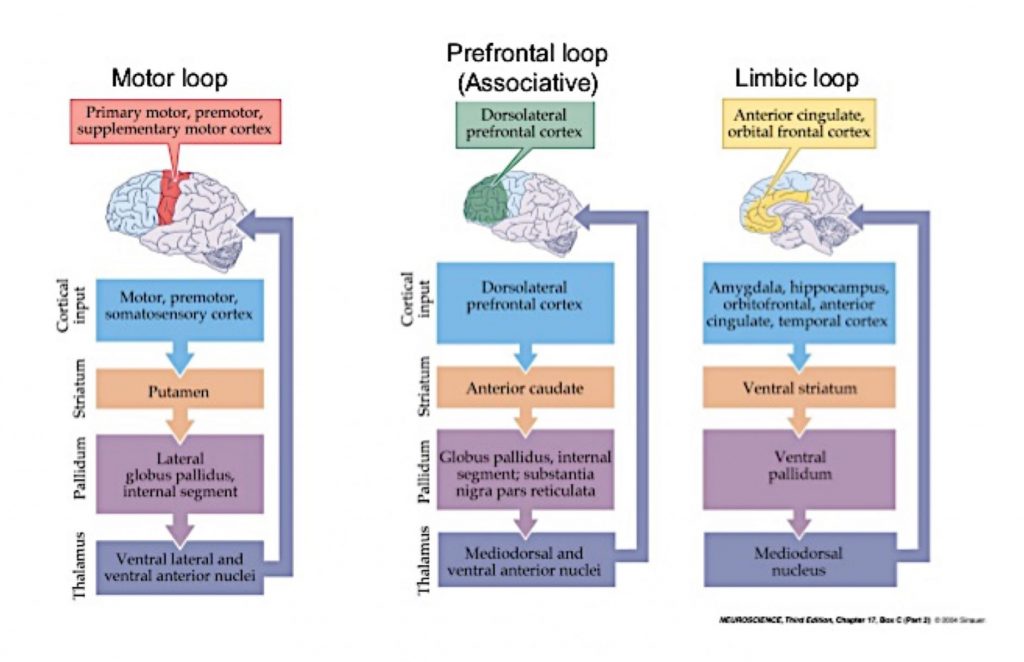
Direct and indirect pathways in the motor loop
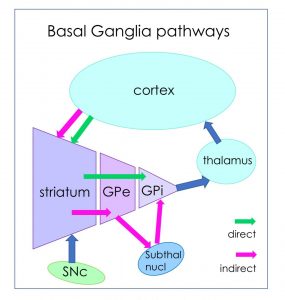
Basal ganglia subcortical pathways
Basal Ganglia Disorders
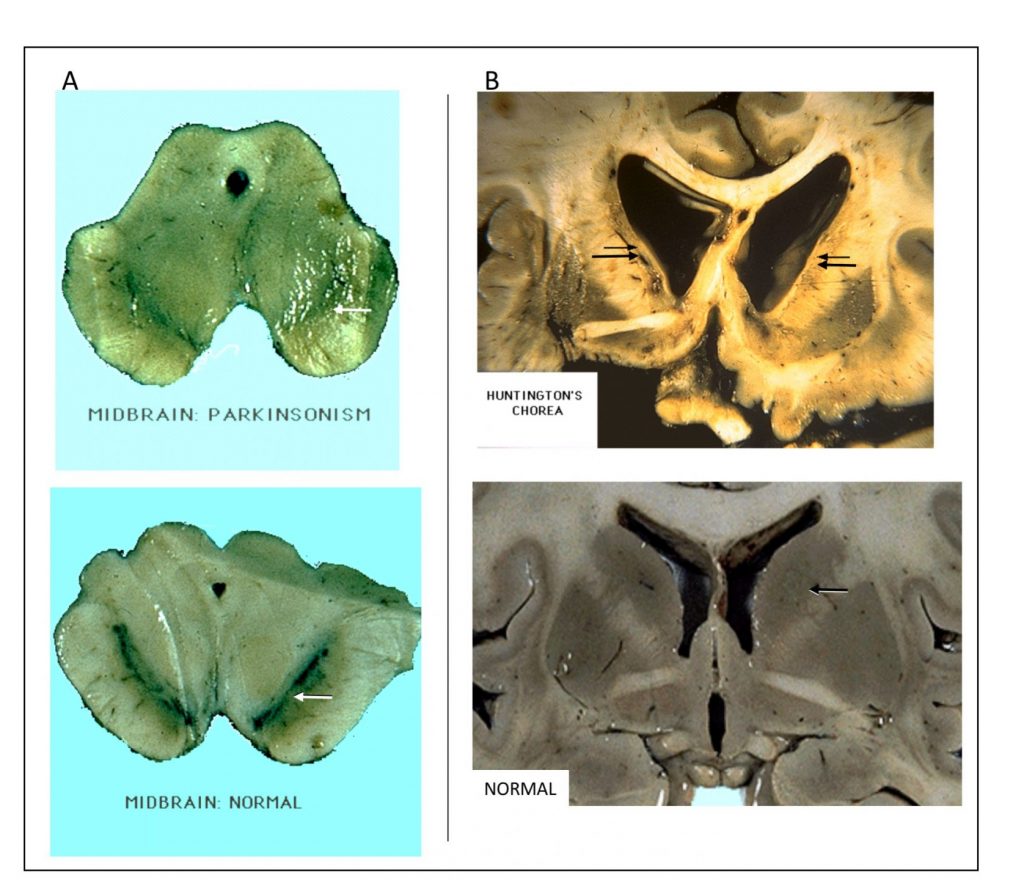
Three important disorders involving the basal ganglia have obvious histopathological correlates (review lecture material for details on 1 and 2):
- Parkinson disease associated with loss of dopamine and decreased staining of the SNc
- Huntington disease associated with marked degeneration of the caudate (leading to abnormally wide anterior horn and body of the lateral ventricle) and putamen.
- Hemiballismus (a movement disorder characterized by wild flailing movement of limbs) is associated with damage to the subthalamic nucleus.
The images below compare sections from pathological specimens (top images) vs normal specimens (bottom images) through the midbrain (left) and forebrain (right).
Arrows indicate regions of interest: A. SNc is diminished in Parkinson disease and B. caudate shows marked reduction and concomitant increase in lateral ventricle profile in late stage Huntington disease.
Cerebellum
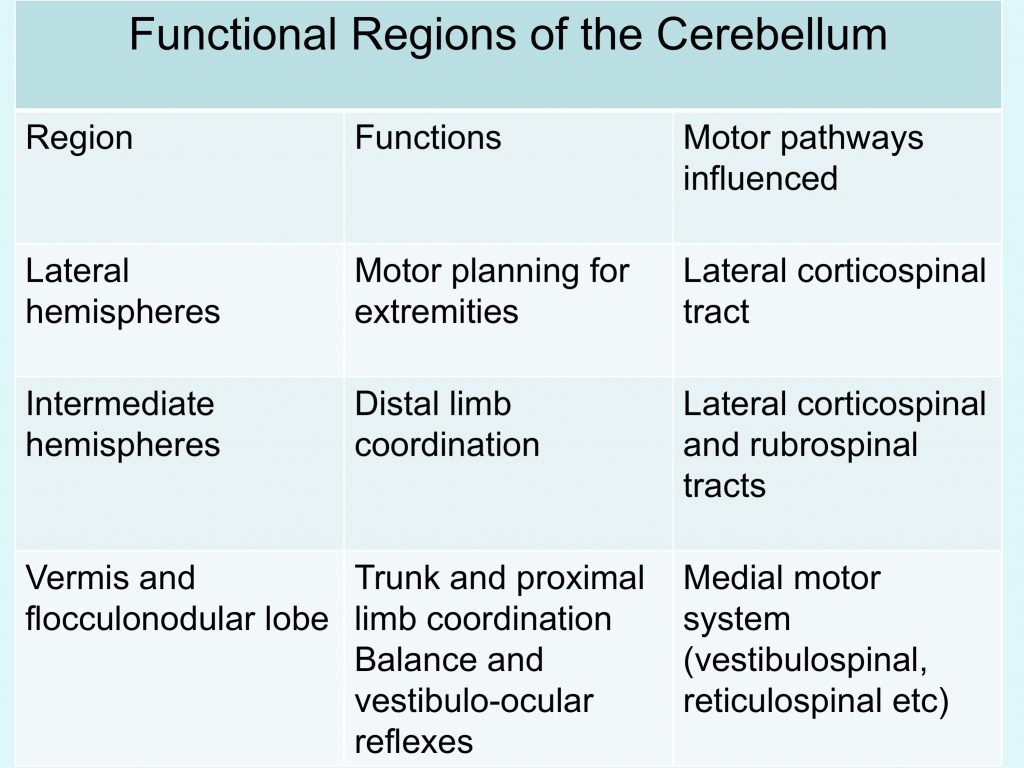
- makes movements fast, accurate, and smooth, serving to coordinate movements that involve simultaneous action at more than one joint or muscle group,
- regulates muscles involved in balance in order to maintain equilibrium and posture
- detects and predicts “motor errors” – the difference between intended movement and that actually performed, both “real-time” and long-term (a type of motor learning).
- plays a role in some cognitive function such as attention related behaviors and language (these roles are not well understood)
Gross anatomy of the cerebellum
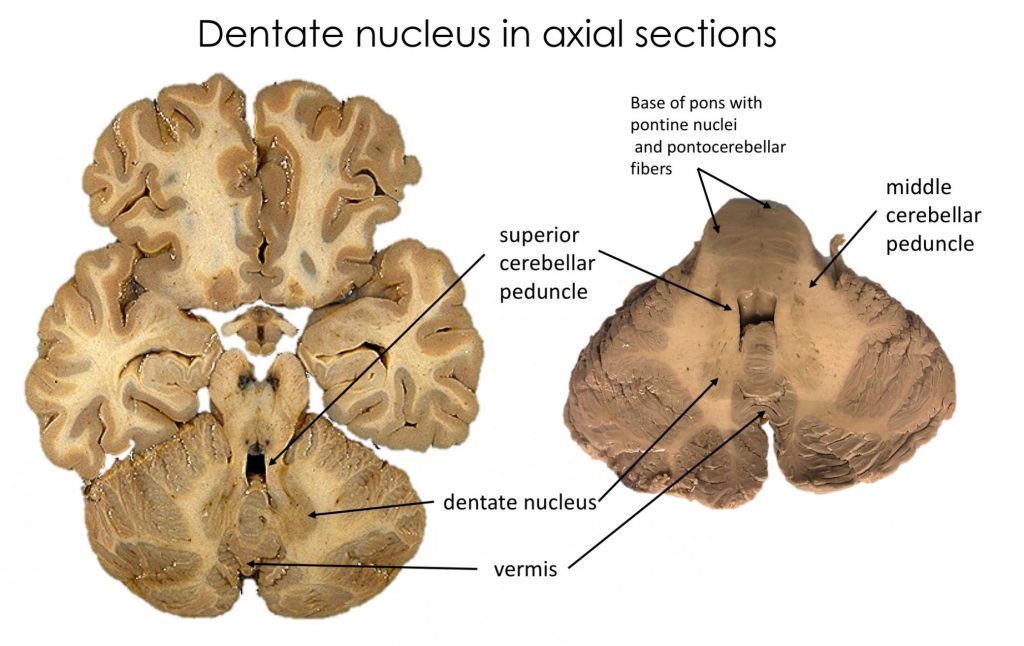
Cerebellar peduncles
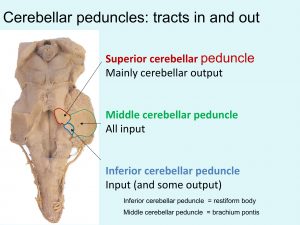
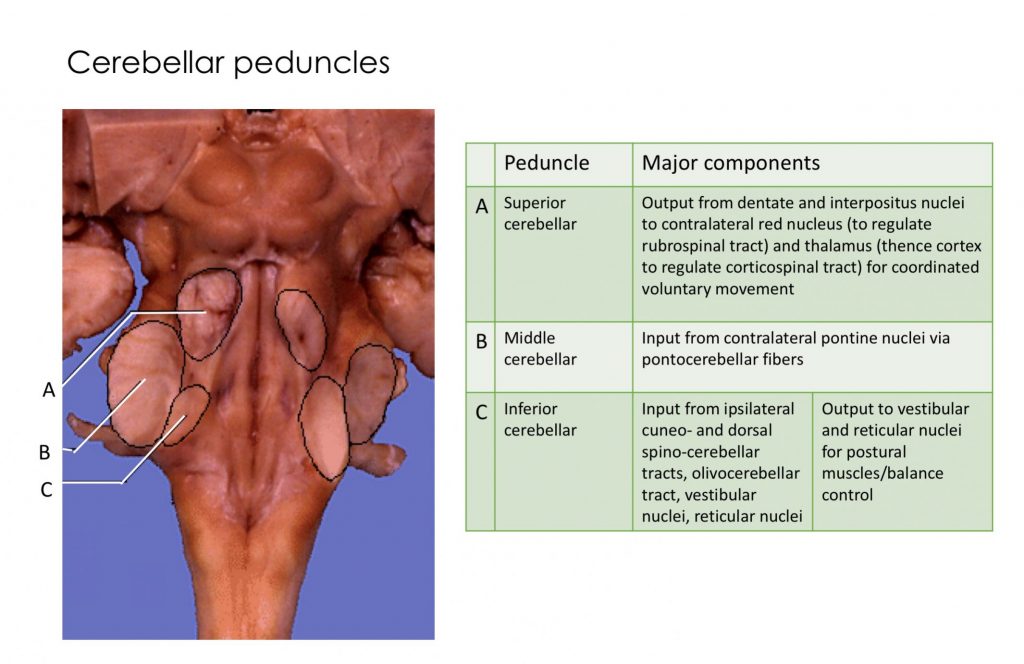
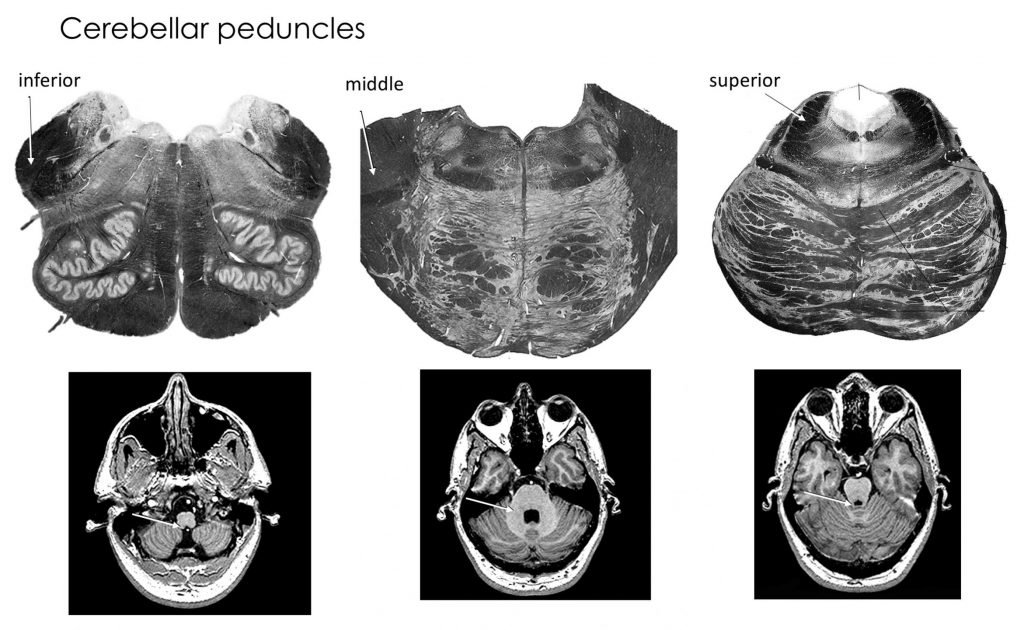
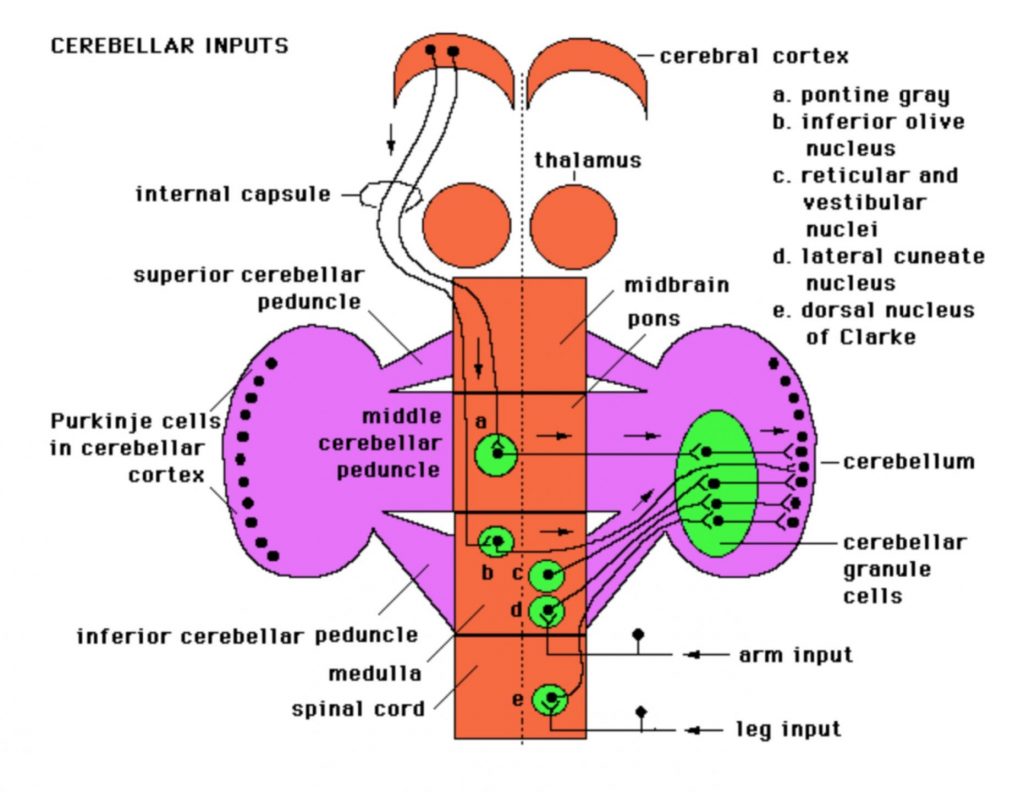
Inputs to cerebellum
- axons of inferior olivary nucleus cells form the climbing fibers, which dominate the dendritic tree of one or at most a few Purkinje cells (this is a unique instance in CNS connectivity)
- axons from all other sources form the so-called mossy fibers which terminate on the granule cells, Granule cells then relay that input to Purkinje via a unique arrangement of axons: each granule cell axon splits into 2 parallel fibers which run transversely, and supply a few synapses to vast numbers of Purkinje cells within a folium.
Practice exercise:
Cerebellar outputs
The most significant output from the cerebellum is via the superior cerebellar peduncle and it crosses the midline to reach the contralateral red nucleus and thalamus (which then relays to cortex). This output is crucial for helping make voluntary movement smooth and coordinated by regulating activity in the corticospinal and rubrospinal tracts.
Other output from the cerebellum (some via the inferior cerebellar peduncle) regulates activity in the vestibulospinal and reticulospinal tracts, thus enabling the cerebellum to influence balance and posture.
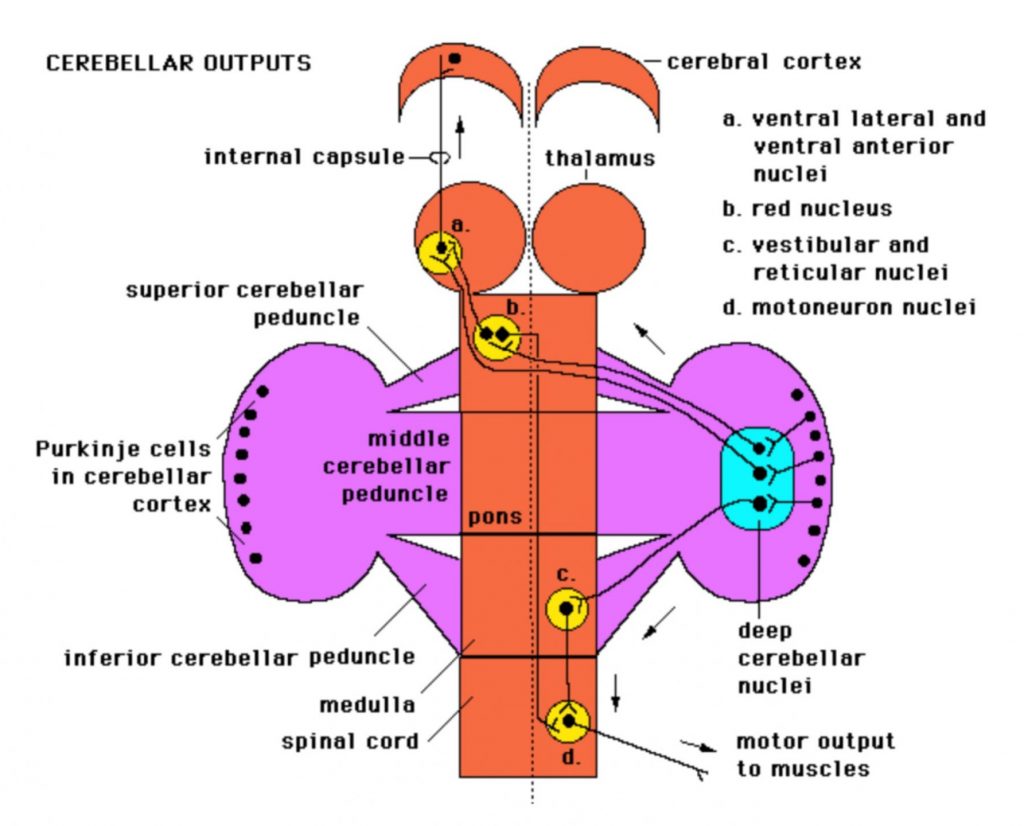
You will note that pathways that interconnect the cortex and cerebellum involve crossing the midline. This makes sense because one side of the cerebellum regulates the same side of the body, while one side of cortex regulates the opposite side. For example to move the right side of the body smoothly and efficiently, the left cortex must communicate with the right cerebellum and vice versa.
Summary of cerebellum function