
8 Lab Week 8: Auditory and Visual Systems
1. Auditory System
Describe the structure of the cochlea, and the mechanism of transduction of auditory stimuli.
Describe the auditory pathway from receptor to auditory cortex.
State the role of the superior olivary nucleus in auditory function.
Locate and identify auditory system structures in gross specimens, histological material and clinical images.
Identify two major parts of cortex involved in language processing: Broca’s and Wernicke’s areas.
Distinguish between conduction hearing loss and sensorineural hearing loss.
2. Visual System
Describe the parts of the eye.
Diagram the basic pathway from receptors to retinal ganglion cells.
State the importance/relevance of the fovea and optic disk.
Locate and identify visual system structures in gross specimens, histological material and clinical images.
Define visual field and explain how the partial crossing of the ganglion cell axons results in half the visual field being represented in the contralateral optic tract, LGN and cortex.
Describe the pathway from the retina to cortex in terms of part of visual field conveyed (retinotopic organization), route of fibers, sites of synapses, and region of termination in calcarine cortex.
Correlate visual field defects with lesion locations in the optic nerve, chiasm, optic radiations, and cortex.
Describe the pupillary light reflex.
The auditory system has evolved tremendous capability to localize and distinguish sound in space and time, forming the foundation of aural communication and language and enriching our lives, for example, with the joy that comes from listening to music. At a basic level, the auditory system codes pitch and amplitude (volume) of sound waves and computes sound location in the environment.
Cochlear Organization
Sound waves are conducted through the external ear and amplified through the action of ossicles spanning the middle ear from the tympanic membrane to the oval window at the base of the cochlea. The cochlea in the inner ear houses the hair cells which are the receptors that transduce sound wave energy.
The cochlea is a bony spiral which contains 3 fluid-filled tubes. The hair cells lie in 1 inner and 3 outer layers on the basilar membrane with their tips embedded in the tectorial membrane. The basilar membrane is taut and narrow at the base of the cochlea and hair cells there respond to high frequencies, while at the apex of the spiral, the basilar membrane is wide and loose and hair cells there respond best to lower frequencies.
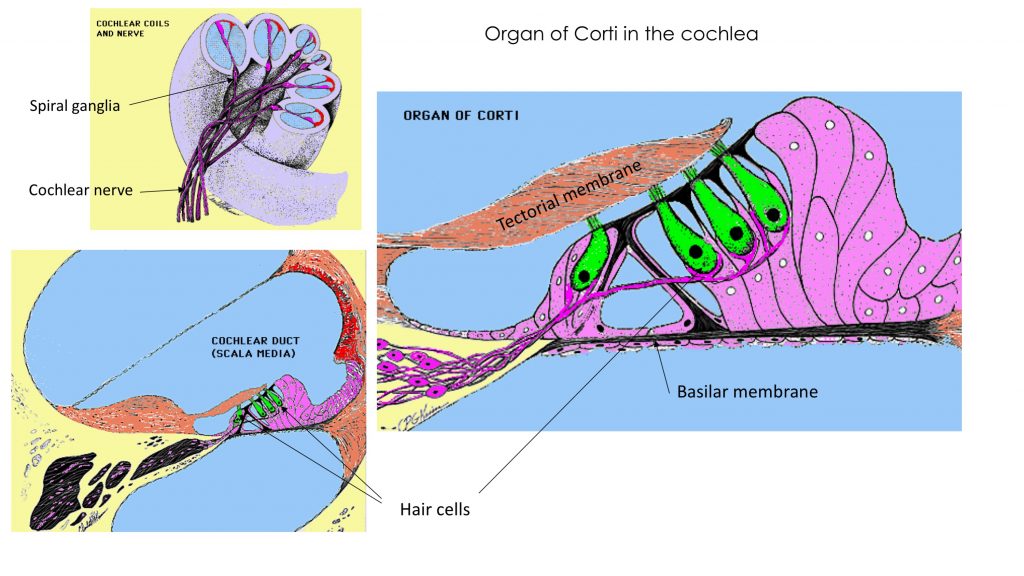
Try this labeling exercise to appreciate the histology of the organ of Corti in the cochlea.
Primary afferent neurons with cell bodies in the spiral ganglia embedded in the bone project through the cochlear part of the vestibulocochlear nerve (CN 8), which attaches to the brainstem laterally at the pons-medulla junction.
The vestibulocochlear nerve (CN 8) carries vestibular information from the semicircular canals for balance, in addition to auditory information from the cochlea.
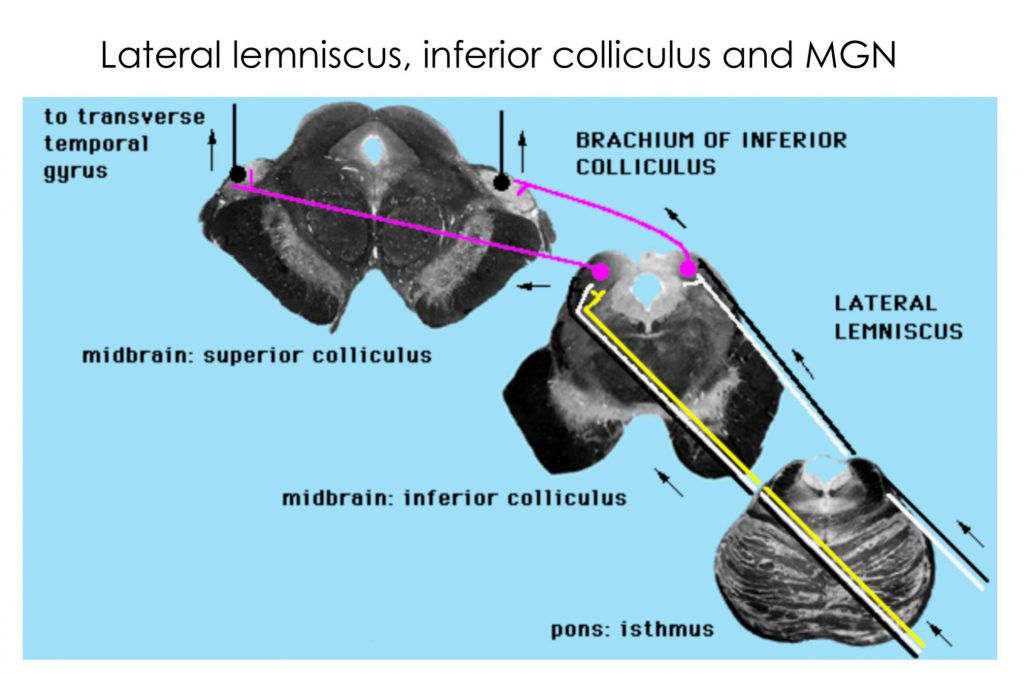
Overview of the auditory pathway
The cochlear fibers in CN 8 send collaterals to the dorsal and ventral cochlear nuclei ipsilaterally. Appreciate that information from one ear is relayed bilaterally through the brainstem, after the first synapse in the cochlear nuclei, i.e., the lateral lemniscus on each side carries fibers from cochlear nuclei on both sides. The lateral lemniscus synapses on neurons in the inferior colliculus, which then project to the medial geniculate nucleus of the thalamus. The medial geniculate nucleus projects to the primary auditory cortex in the transverse temporal gyrus via the auditory radiation, a part of the internal capsule that is not easily visible.
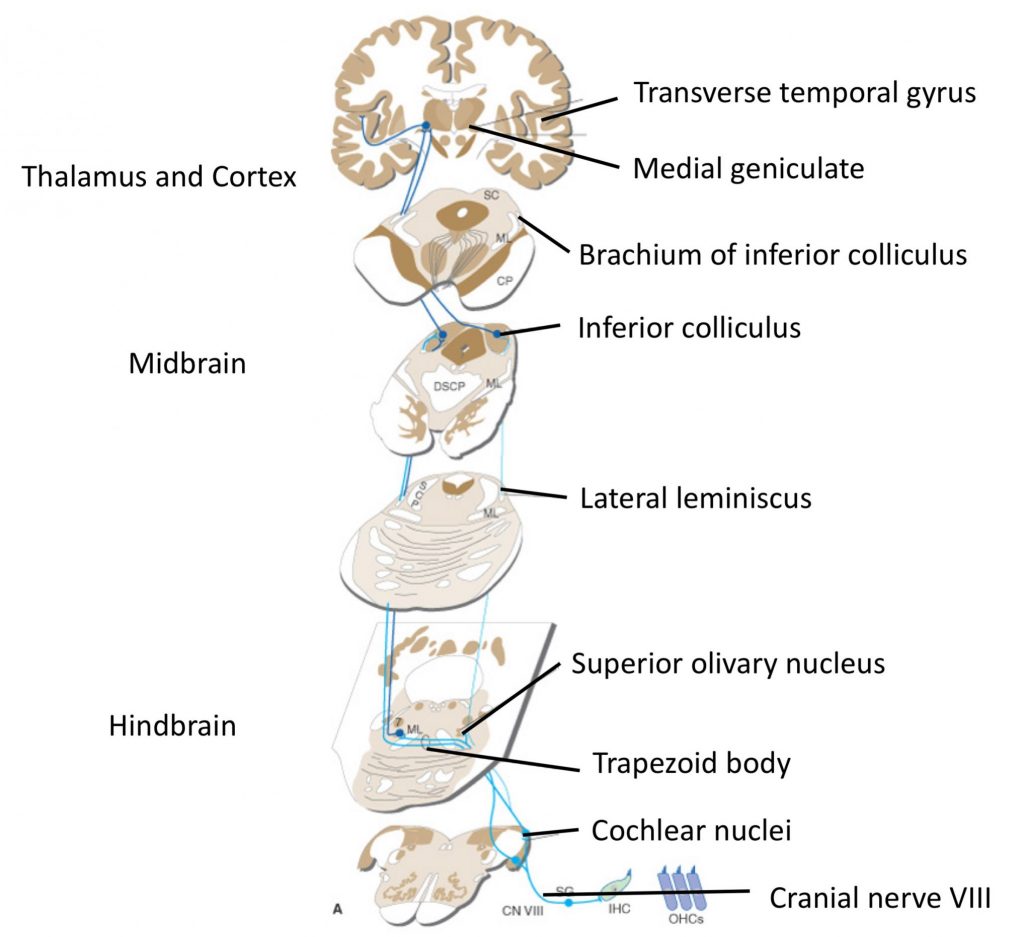
There are many more nuclei in the brainstem that process auditory information; one shown in the diagram above is the superior olivary nucleus, which processes inputs from both ears to determine sound location, on the basis of differences in timing and intensity of sound at each ear. Also note there are crossing fibers carrying auditory information at many levels in the brainstem. The auditory pathway is thus binaural and from the cochlear nuclei rostrally, damage to the pathway on one side does not result in loss of hearing in just one ear.
This movie shows a computer graphic reconstruction of the basic pathway.
Study the following sections through brainstem to familiarize yourselves with auditory pathway structures.
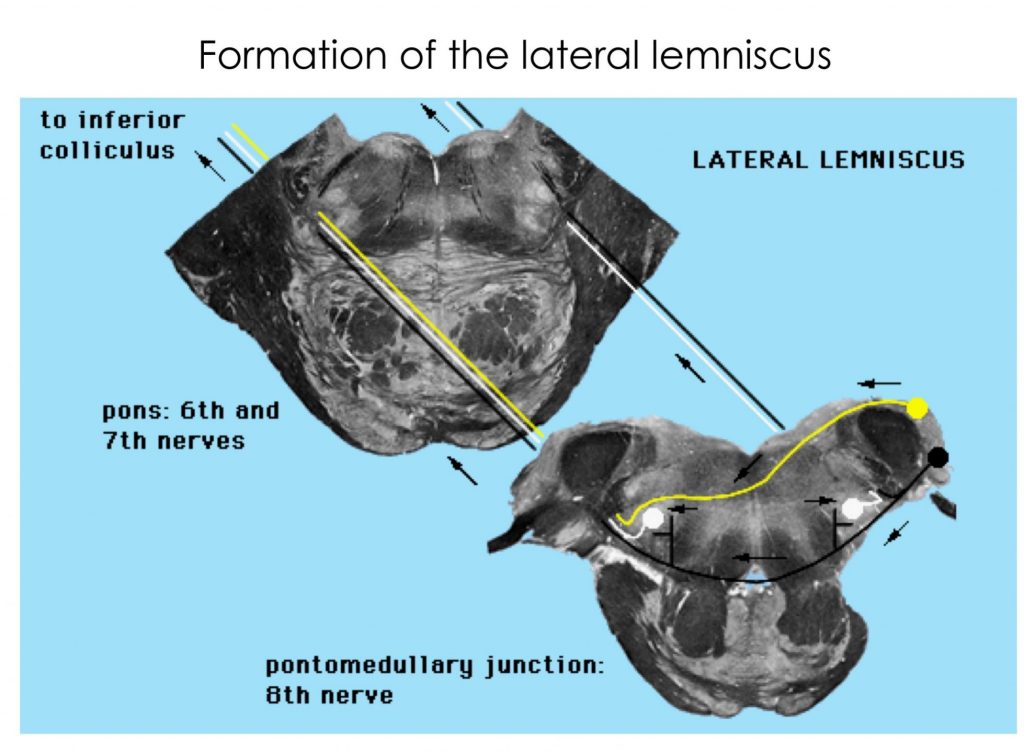
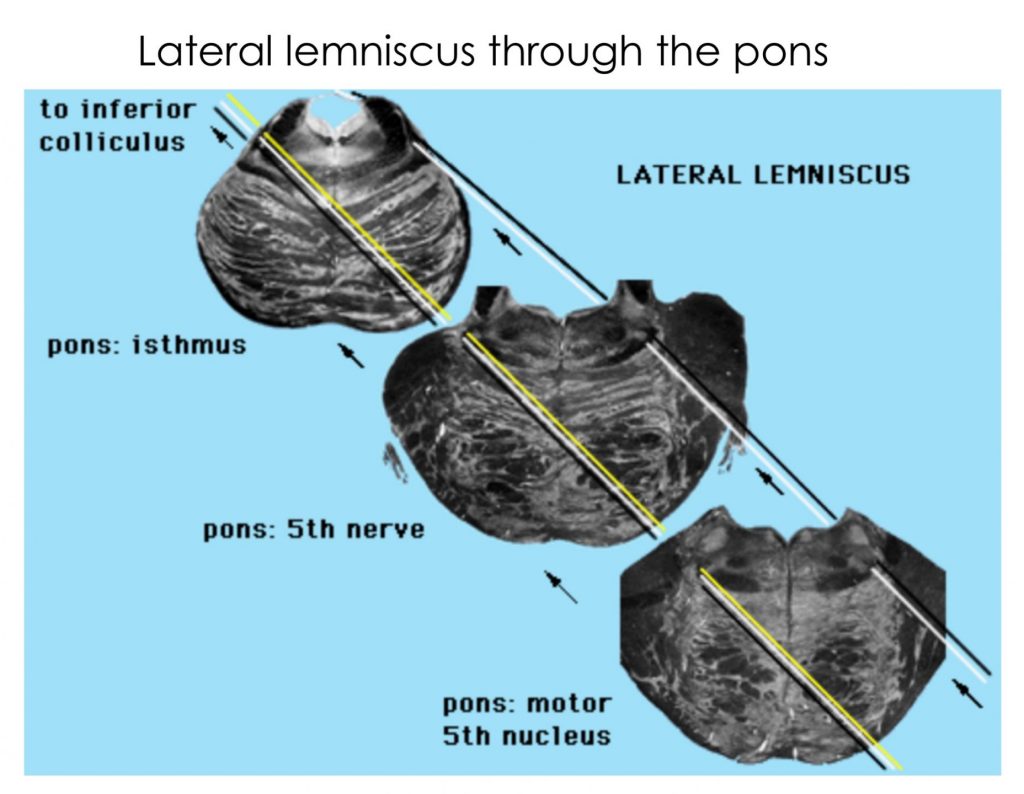
Try these quizzes!
Auditory cortex
The first cortical processing center for audition (primary auditory cortex, A1) is located within the transverse temporal gyrus. It is not possible to see this on an intact brain specimen because it sits obliquely in the superior temporal gyrus, buried in the lateral fissure. The image below shows a specially dissected preparation on the left. Use the full screen option and examine the structures seen in axial and frontal planes that capture the transverse temporal gyrus.
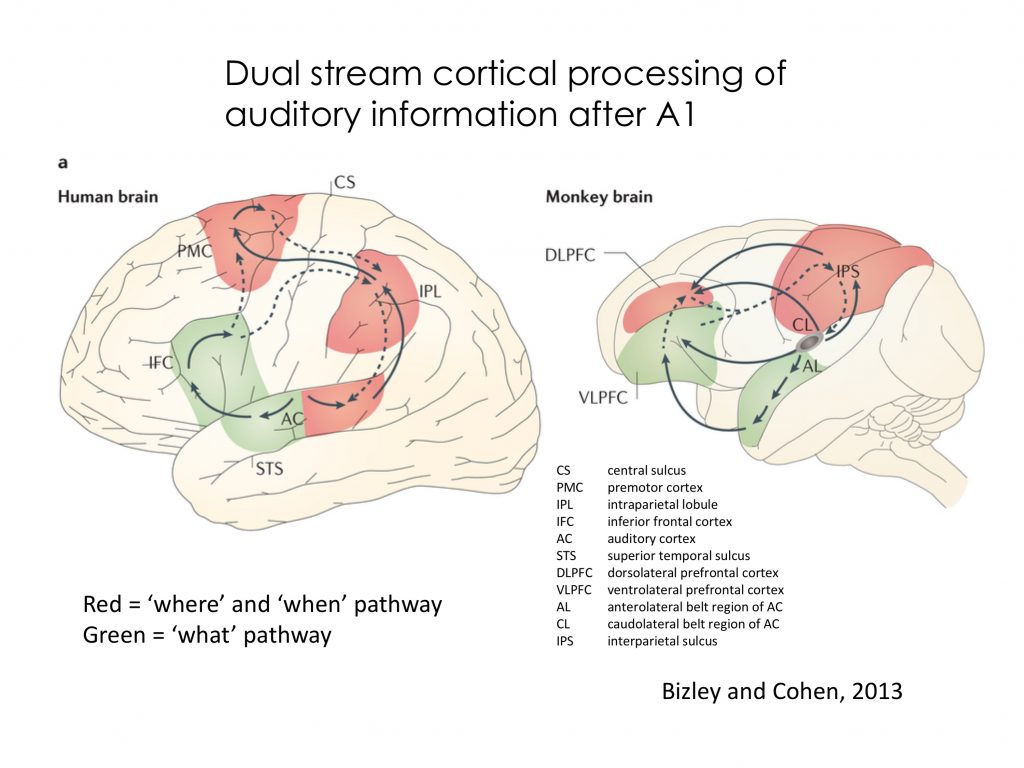
From primary auditory cortex, in analogy to the visual system, dual pathways have been proposed through higher auditory cortex for “where and when” processing (parietal cortex) and “what” processing (anterior and infratemporal cortex) as shown in the image below (this is just for enrichment!)
Hearing Loss
Because the auditory pathway through the brainstem is binaural, unilateral hearing loss is generally due to damage at locations distal (peripheral) to the cochlear nuclei.
Hearing loss can be classified as conduction or sensorineural hearing loss, depending on the structure most impaired. Conduction hearing loss refers to loss related to abnormalities in the conduction of pressure waves to and through the external and middle ears, for example, due to physical blockage in the auditory canal, perforation of the tympanic membrane, or sclerosis of the ossicles. Sensorineural hearing loss is due to damage to the receptors or the neural pathways conveying auditory information. Examples include hair cell damage due to ototoxic chemicals or loud noises, or acoustic neuromas (more correctly also known as vestibular schwannomas) that compress the vestibulocochlear nerve.
Language Areas
There are two main areas related to language: Broca’s area is the “motor or expressive” language area located in the inferior frontal gyrus, and Wernicke’s area is the “sensory or receptive language” area located around the posterior end of the lateral fissure. These areas are interconnected by a bundle of fibers (the arcuate fasciculus) that runs in an arc between temporal, parietal and frontal lobes. Disruption of either area or this bundle of interconnecting fibers leads to language problems (aphasias). The equivalent areas in the non-dominant hemisphere are responsible for “emotional” aspects of language, e.g. the inferior frontal gyrus in the non-dominant hemisphere plays a role in adding meaningful intonation to speech (prosody), while the region matching Wernicke’s area appears to play a role in decoding the meaning of intonation, e.g., in sarcasm, anger, or questions.
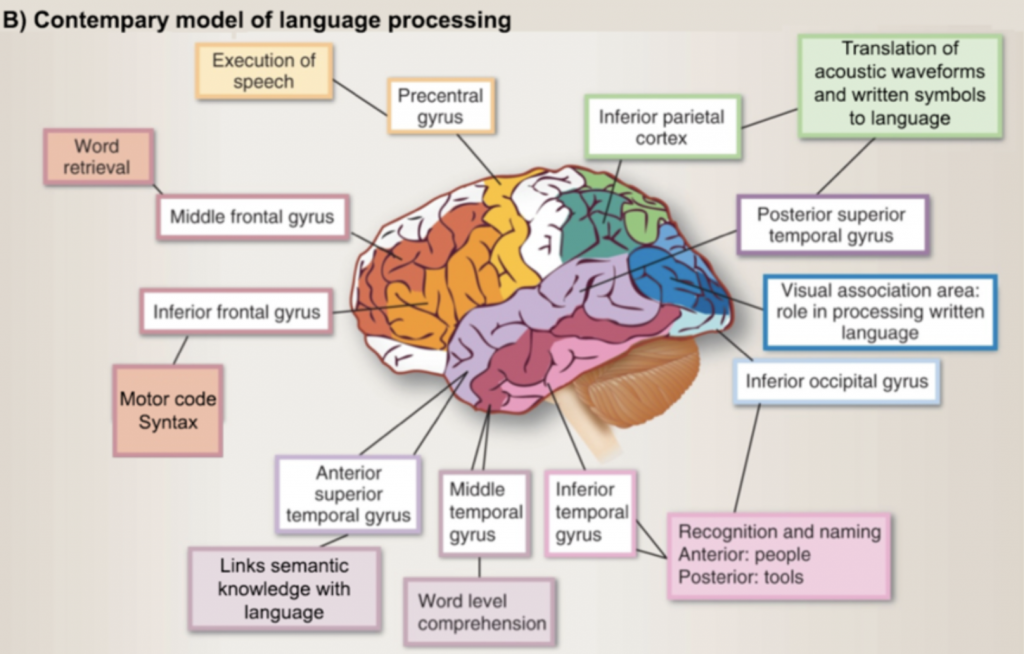
Of course, language in the brain is far more complex than this simplistic view. Some estimates suggest that language processing occurs in upwards of 60% of our cortex, forms the foundational neural code supporting many forms of higher cognitive functions, and generates a vast array of skills that make us uniquely human (see shaded ares in image below–again, this is purely for enrichment!).
Summary
Watch this video from Dr. Stensaas at University of Utah. It is helpful to see the actual sections through the brainstem (around minute 13) that she shows after a demonstration of models of the ear.
Visual System
Visual information is used in perceiving the environment, in controlling the pupil, and in regulating circadian rhythms. Receptors in the eye transduce light and the optic nerve and tract carry that information to a variety of targets.
Primary Visual Pathway
The structures that make up the primary visual pathway for conscious perception are located relatively close to the surface of the brain and can be easily dissected as shown in the image below.
Retinal ganglion cell axons leave the eye at the optic disk to become the optic nerve (CN 2). Axons from nasal retinal ganglion cells cross at the optic chiasm, while those from the temporal retinal ganglion cells pass into the ipsilateral optic tract. This partial crossing at the chiasm brings together all the information from one visual field to be processed by the opposite thalamus and cortex.
The vast majority of axons in the optic tract synapse in the lateral geniculate nucleus (LGN) of the thalamus, which in turn projects through direct radiations and through Meyer’s loop to the primary visual cortex in the banks of the calcarine fissure. This is the pathway that underlies conscious visual perception.
Study the image and click on the “i” symbols for more information.
Other targets of visual input
This image highlights targets of retinal ganglion cell axons.
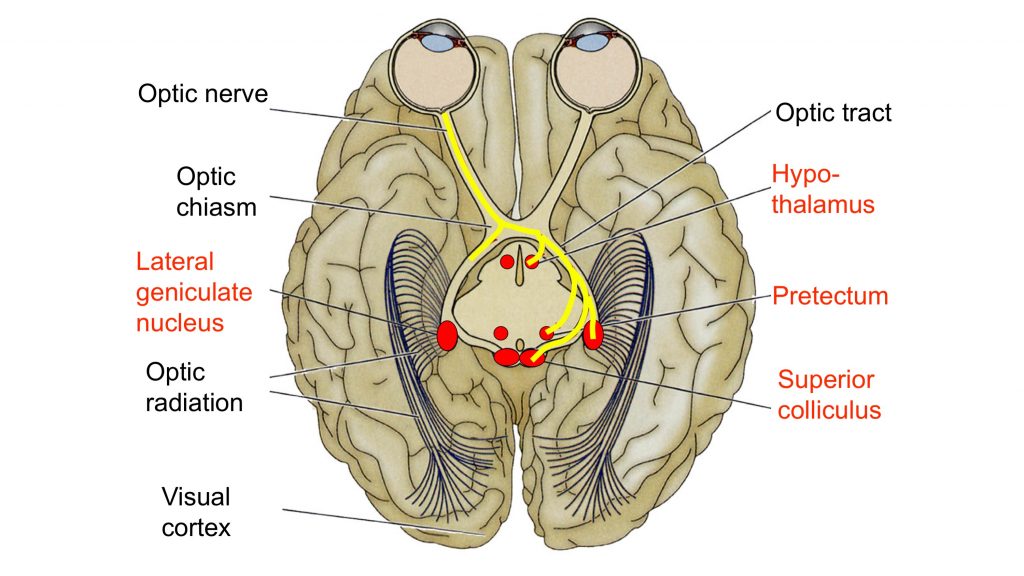
Some retinal ganglion cells axons peel off from the optic tract and enter the tectum of the midbrain where they terminate in the superior colliculus. The superior colliculus is associated with controlling gaze shifts and saccades. A saccade is a rapid eye movement that directs gaze to an object of interest, ie, the eye movement serves to foveate or project the image of the object onto the foveal region where acuity is best.
Another target of some of the axons in the optic tract is the pretectal area, a small region at the boundary between the LGN and the superior colliculus. This nucleus in the midbrain just anterior to the superior colliculus (see lab atlas, S12, S13 & H4) projects to the Edinger-Westphal nucleus (located adjacent to the oculomotor nucleus), which sends parasympathetic output in CN 3 to the iris and causes pupil constriction in the pupillary light reflex (see below).
The suprachiasmatic nucleus (SCN) is yet another target for retinal ganglion cell axons. This nucleus, a small structure located just above the optic chiasm in the anterior hypothalamus, serves as the master circadian clock for the body and brain. The “circadian rhythm” is an approximate 24-hour oscillation seen in many physiological processes such as hunger and, importantly, the sleep/wake cycle. Intrinsically photosensitive retinal ganglion cells that express the opsin-like protein melanopsin project to the SCN.
Eye and Retina
Structures of the anterior eye help control the amount of light entering the eye and refraction of light onto the retina. An important fact to remember is that the lens inverts and reverses an image as it is projected onto the retina.
An elegant and complex system coordinates movements of the 2 eyes through control of 3 pairs of cranial nerve nuclei. Sensory information from the eyeball itself, carrying for example, touch, pain, and pressure information, is conveyed via the ophthalmic division of the trigeminal nerve. Thus a total of 5 of the 12 cranial nerves (CN 2, 3, 4, 5, and 6) play a role in eye and visual function.
Recall that the neural components of the eye (retina and optic nerve) are derived from the central nervous system, so the retina contains not just the photoreceptors that transduce light, but also contains the complicated circuitry that begins to process visual information before the axons of the retinal ganglion cells leave the eyeball to form the optic nerve.
Review the layers of the retina and label them here. Recall that this initial part of the visual pathway is photoreceptor⇒bipolar cell⇒retinal ganglion cell.
In the fovea, the overlying layers are “pushed to the side” to allow better access of light to photoreceptors as seen here. This image, a cross section through the retina and choroid of a macaque monkey, shows the pigment epithelium, choroid and scleral layers nicely as well as the foveal “pit”.
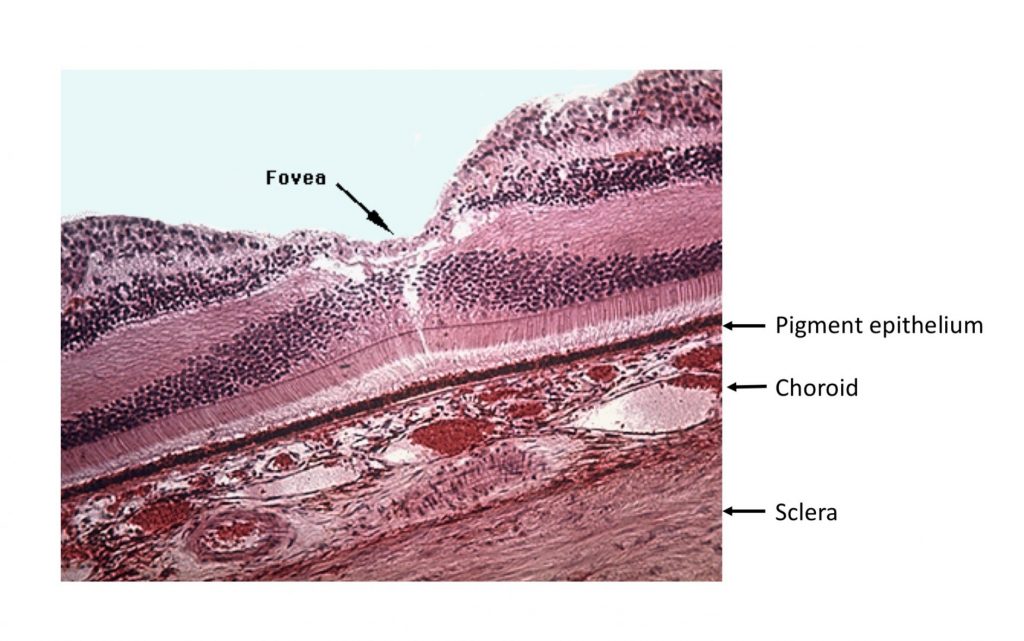 Recall that the fovea is specialized for high acuity and color vision, and has the highest density of cones, and no rods.
Recall that the fovea is specialized for high acuity and color vision, and has the highest density of cones, and no rods.
Fundoscopic views of the retina show another specialization of the fovea–blood vessels surround it but do not pass across it. Also note in this view the appearance of the optic disk where the ganglion cell axons accumulate and exit the eye to form the optic nerve. The retinal blood vessels enter and exit through the center of the disk. Because there are no photoreceptors in the disk, there is a blind spot in the visual field that can be demonstrated under certain conditions. Viewing the optic disk with an ophthalmoscope can also be a way to assess when intracranial pressure is elevated: the edges of the disk then appear blurry.
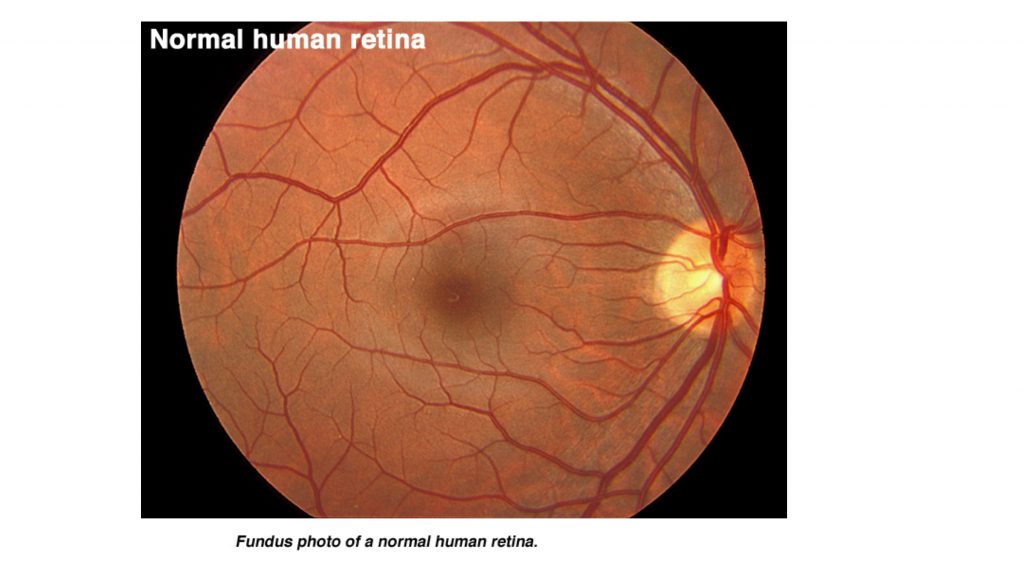
The optic nerve continues as the optic tract beyond the chiasm and passes laterally to synapse in the LGN.
Find the optic nerves, chiasm, tract and visual cortex in these images. These exercises have “hidden hotspots” that will show when you click on them.
Visual Field
The concept of visual field is crucial for understanding visual field deficits. Take a moment to map out your own visual field.
- Fixate on a point in front of you with both eyes open. Realize that a vertical line drawn through that point represents the division between the left and right visual fields (not the left and right eye fields)
- Maintain your fixation. But close the right eye, and move a finger of your right hand from out of the field of view on the right in towards the midline in a horizontal plane. Note that the point at which you first see the finger is well to the right of the vertical line dividing left and right fields. So the left eye sees the left field and part of the right field.
- Repeat with the left side. You can thus establish the binocular part of the visual field. Because of this overlap, each eye needs to be tested individually to determine field loss.
- Realize that the lens inverts and reverses the image, so that temporal field is projected on the nasal retina and vice versa, and the upper field is projected onto the lower retina. In clinical practice, you will generally be describing the field deficit not the retinal projection.
- The key point to remember: left field is projected on the right sides of each retina and vice versa. The partial crossing (of just nasal retinal ganglion cell axons) at the chiasm allows all information from one half of the visual field to be processed in the opposite side of the brain from the optic tract onwards. So, lesions retrochiasmatically result in some visual loss of the contralateral field, manifested as a loss in each eye.
- The convention for presenting visual field testing is as though you were the patient looking outwards, left eye on left, right eye on right, shading usually represents the loss of vision.
Topographic representation and visual fields
The primary visual pathway is retinotopically organized. View this video to see how the visual field is mapped through the system and onto primary visual cortex. This mapping allows for prediction of deficits with lesions to specific parts of the visual pathway.
One fact not mentioned in the video is that the most central field is represented at the most caudal pole of the occipital lobe, and the central to peripheral axis is mapped posterior to anterior along the calcarine sulcus. Lower field is represented in part of the upper bank of the calcarine sulcus and the upper field in the lower bank. Remember to look for the interactive exercises represented as circles in the video timeline.
Try the exercise below to confirm your understanding of the field representations.
Higher order visual processing and Visual processing streams
The complex web of connectivity between primary visual cortex and higher order cortex supports the concept of a visual processing hierarchy: simple visual features such as lines, orientation and simple shapes are processed early within the hierarchy whereas regions further along the pathway (more synapses away from V1, primary visual cortex) process more complex visual stimuli, relying on input about various simple features of a visual stimulus from early areas to construct more complex representations and analysis of visual objects and scenes.
Two well established processing streams, the dorsal and ventral streams, begin in V1. The ventral stream continues as a multisynaptic pathway ventrally terminating within the middle and inferior temporal lobe; it is referred to as the ‘what’ pathway and plays a role in object identification based on shape, size, form and orientation. Within the ventral processing pathway higher-order visual regions exist that are sensitive to various complex visual stimuli, eg. the fusiform gyrus contains a “face area” where cells show sensitivity to images of faces. An additional inferotemporal region, the parahippocampal place area, is sensitive to environmental scenes. For example, this area responds to topographical scene stimuli such as forests, landscapes, cityscapes, or rooms (the general idea being ‘images of places’).
The dorsal or ‘where’ (and how) pathway courses dorsally ending in the posterior parietal cortex. It relays information about object location to the parietal lobe, in part for visuomotor integration.
Pupillary light reflex
The pupillary light reflex serves to adjust the pupil size to environmental light conditions to optimize visual performance. At low light levels the pupil dilates to a maximal diameter. With increasing light level the pupil constricts to reduce the total light impinging on the retina. The pupillary light reflex is one of many neural mechanisms that allow our visual system to adapt, ie., adjust its sensitivity, to light levels that can vary by up to 12 log units from full outdoor sunlight to moonless starlight.
Normally, both pupils constrict when light falls on just one eye. Two factors contribute to bilateral pupil constriction: first, axons from one eye go to both the left and right pretectal nuclei, and second, the left and right pretectal nuclei are connected to both Edinger-Westphal nuclei. If light shone in one eye fails to cause any constriction of the contralateral pupil, a sensory explanation is a prechiasmatic lesion (eg of the optic nerve), because damage central to the optic chiasm would leave the pathway to the opposite eye at least partially intact. In contrast, if CN III is damaged, the affected eye’s pupil remains dilated no matter which eye is stimulated with light, because the parasympathetic motor pathway controlling the iris has been lost.
The pupillary light reflex components are shown below: Left, top-down view of visual pathway to pretectal nuclei, and output back to iris. Right panel, consequences of damage to the right optic nerve on pupil size. A: dim light, moderately dilated pupils; B: light in left eye causes both pupils to constrict. C: light in right eye causes no pupil constriction.
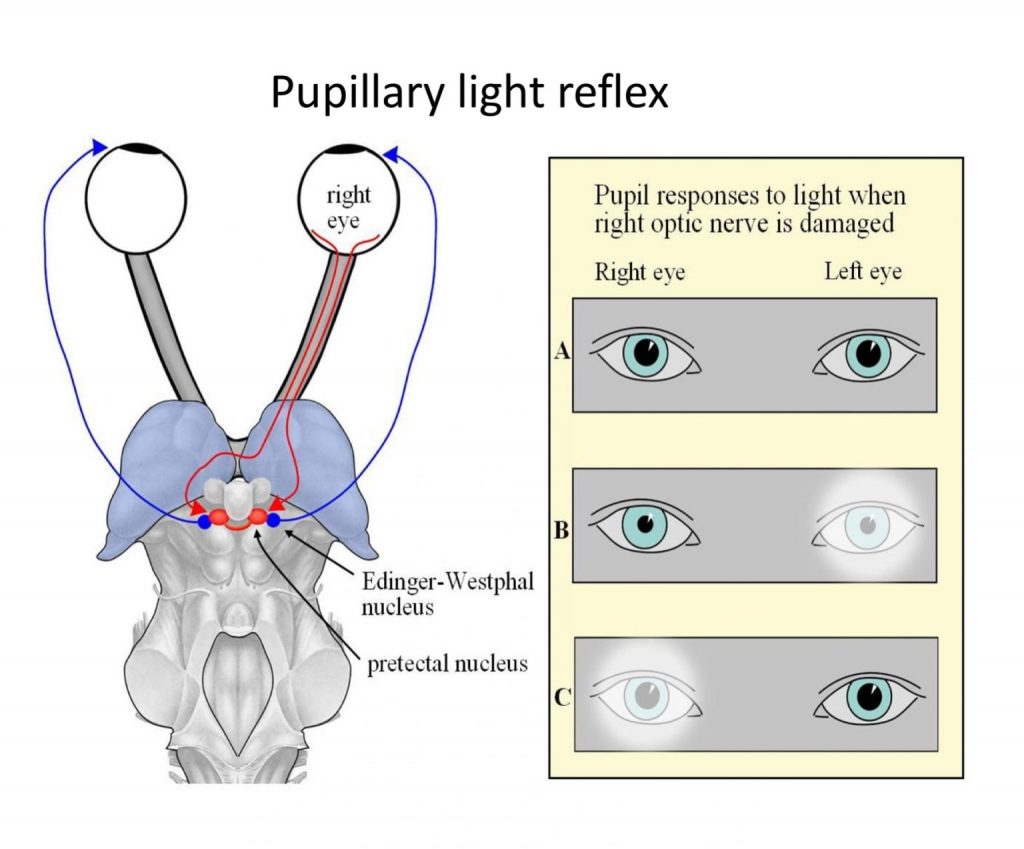
Pupils normally dilate when light levels are reduced. This dilation occurs due to activity in the sympathetic system, and involves a pathway that originates from the upper thoracic spinal cord.
Structures to Identify