
Muscle Tissue
SESSION OBJECTIVES
- Be able to distinguish the three types of muscle.
- Examine examples of striated muscle.
- Review the basis for the striations and mechanism of contraction.
- Examine a neuromuscular junction.
- Review the mechanism of synaptic communication and motor units.
- Examine smooth muscle in various contexts and distinguish from connective tissue.
- Identify the distinguishing characteristics of each muscle type that relate to their unique properties.
- Compare and contrast muscle mechanics and energy use in the three muscle types
OPTIONAL PRE-CLASS MATERIALS FOR THIS SESSION:
- Skim the section titles, bolded terms, and image captions from Junqueira’s Basic Histology, Chapter 10 (Click here for FMR library resources) to fill in any knowledge gaps you feel you have.
OVERVIEW:
There are three basic types of muscle:
- Skeletal muscle is required for obtaining food, eating, drinking, breathing, moving and reproducing.
- Cardiac muscle is required for moving blood cells to all tissues of the body.
- Smooth muscle is required for maintaining blood pressure, for digestion, breathing, excretion and parturition.
Movements are produced in all three types of cells by the contractile apparatus, which is a highly specialized cytoskeleton, called the sarcomere. Each cell type has its own brand of myosin, actin, and other proteins in the contractile apparatus.
SKELETAL MUSCLE (Session Objectives #1, #2, #3, #6)
Knowledge check: Mentally identify each structure before clicking on the question mark to check your answer:
- Skeletal muscle can regenerate from satellite cells, reserve stem cells adjacent to the muscle fiber called myosatellite cells.
- Muscles are made up of many long and thin cells and each individual cell is called a muscle fiber.
- Muscle fibers are grouped together into fascicles; endomysium surrounds and occupies the space between the fibers;
- Fascicles are surrounded by perimysium;
- The entire muscle is surrounded by epimysium;
- Blood vessels and lymphatic vessels lie within the perimysium and endomysium
- Muscle fibers are multinucleate cells, formed by fusion of many individual myoblasts during development. In mature muscle fibers, the nuclei sit just underneath the cell membrane (called the sarcolemma).
- Within each cell the cytoskeleton is organized into many long cylindrical myofibrils that extend the length of the muscle cell.
- Each myofibril is a bundle of F-actin (thin) and myosin (thick) filaments organized as repeating units called sarcomeres.
- The sarcomere is the functional unit of skeletal and cardiac muscle.
- The repeating unit of the sarcomere is what gives the muscle fiber its striations, and sometimes skeletal muscle is called striated muscle.
- A sarcomere is bounded on either side by Z-discs (aka Z-bands). Actin filaments extend from the Z-discs towards the center of the sarcomere on either end; myosin thick filaments, located in the center of the sarcomere, form cross-links with the actin filaments via their “head” domains. The actin and myosin filaments are interdigitated.
- The overlap between the thick and thin filaments leads to different patterns of birefringence and these zones have names: proceeding from the center of the sarcomere, the M, H, A, I and Z bands or zones. It is important for you to know how these bands relate to the proteins of the sarcomere. For example, the A-band demarcates the length of the thick filaments, and so it appears darker in most images. By contrast, the I-band represents the thin filaments, between their attachments to the Z-discs to their overlap with the thick filaments.
Knowledge check: Identify the structures of the sarcomere viewed by electron microscopy and answer the following questions:
During contraction, the thick and thin filaments slide past one another in this ratcheting process, shortening the length of each sarcomere and consequently the length of the muscle fiber. Your text describes this process very well. You should also know what happens to the length of the various bands during contraction: for example, the I-bands will get shorter during contraction with the increasing overlap between thick and thin filaments.
- Summary of the cross-bridge cycle of skeletal muscle contraction:
ATP and contraction: after the myosin head has completed a power stroke, ATP is required to break the actin-myosin links to allow another power stroke. The myosin heads have an ATPase, an enzyme that hydrolyzes the ATP to ADP and uses this energy to break the links. Without ATP, the bonds between actin and myosin are stable; rigor mortis occurs in muscles as a result of the lack of ATP at death.
- Calcium and muscle contraction
- The sarcoplasmic reticulum is the smooth ER of the muscle fiber;
- The sarcoplasmic reticulum sequesters Ca+2 ions and releases them in response to depolarization of the muscle fiber membrane.
- The depolarization of the muscle fiber membrane is communicated to the sarcoplasmic reticulum via finger-like invaginations of the surface membrane, called T-tubules.
- The connection between the T-tubule and the sarcoplasmic reticulum is called a triad.
- Calcium and muscle contraction: When the membrane of the muscle fiber is depolarized by the nerve (see below), causing release of Ca+2 ions from the sarcoplasmic reticulum, the Ca+2 ions bind to a protein called troponinC (which is part of the thin filament) and causes a conformational change in the troponin to expose myosin binding sites on the actin filaments. The myosin heads start the ratcheting process described above as long as the Ca+2 ions are around, but when the muscle fiber membrane depolarization ends (i.e., no more nerve stimulation), the Ca+2 ions are again sequestered into the sarcoplasmic reticulum.
- The neuromuscular junction (NMJ; Session Objective #4) is the synapse between the axon terminal and muscle fiber (sometimes called the motor endplate).
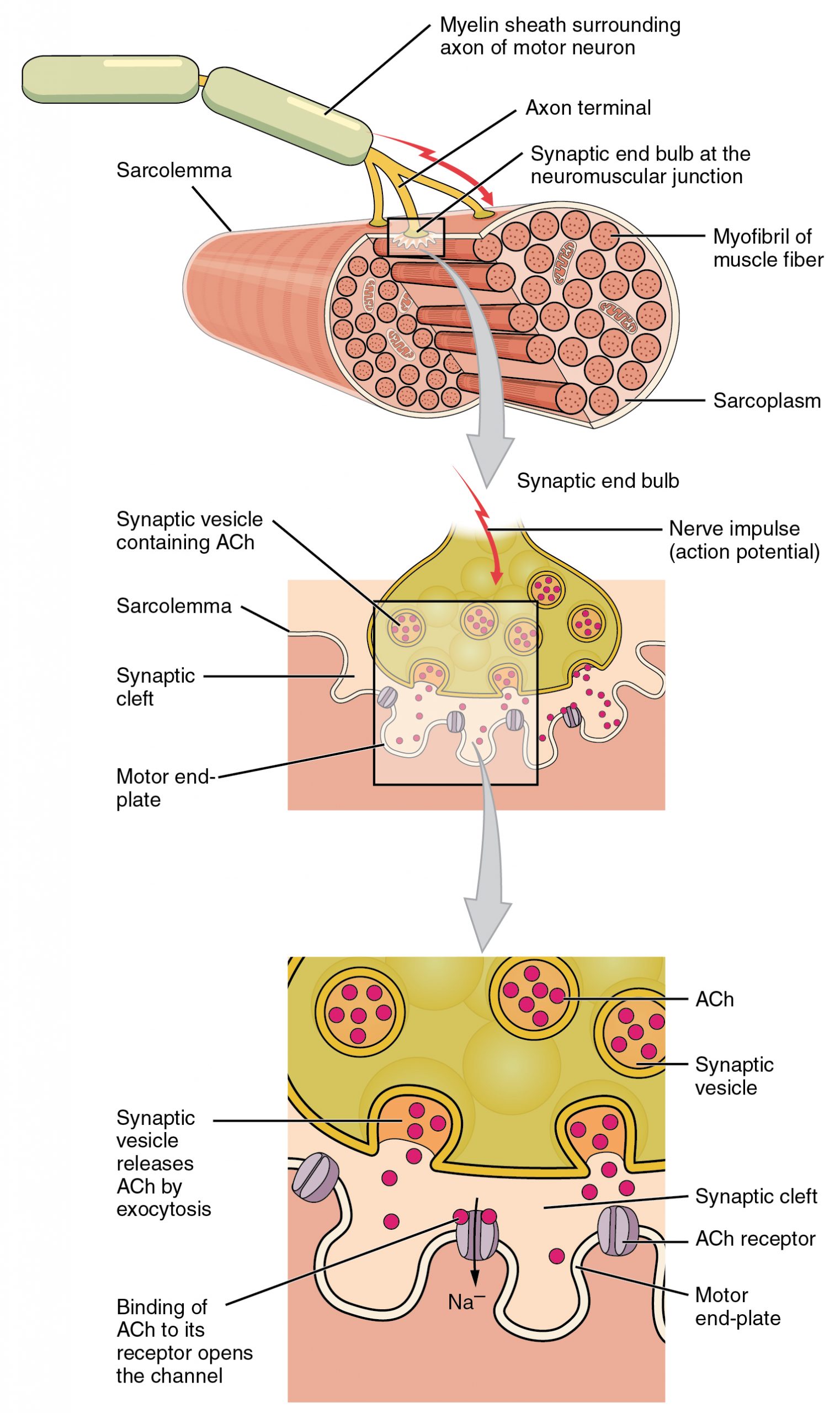
1. Presynaptic specializations. Like other synapses, the terminal of the axon of the motor neuron has clusters of synaptic vesicles and active zones for release adjacent to the junctional folds. Many of the same proteins are involved in the release of the vesicles as in other synapses. Botulinum toxin, BOTOX, inhibits muscle contraction by inhibiting the proteins required for vesicle fusion.
2. Postsynaptic specializations: Like other chemical synapses, the NMJ has neurotransmitter receptors clustered in the postsynaptic membrane; however, in the NMJ the muscle fiber membrane is highly folded into a series of junctional folds; the acetylcholine receptors are clustered near the tops of the folds. Myasthenia gravis is an autoimmune disease that leads to weakness and paralysis because the acetylcholine receptors are inhibited by antibodies.
3. A single NMJ has multiple neurotransmitter release sites.
4. There is a thin extracellular matrix in the synaptic cleft that remains after the nerve is damaged and allows the nerve to find the correct site to reform the NMJ when it regenerates.
5. The axon from a single motor neuron branches at its terminal arbor and can innervate many muscle fibers to form a motor unit. All the muscle fibers in the motor unit contract together when the motor neuron is activated.
- Sense organs in muscles and tendons enable the nervous system to monitor muscle stretch and tension to maintain tone.
- Muscle spindle:
- Location: Within the muscles
- Structure: Connective tissue capsule surrounding a few non-striated intrafusal muscle fibers
- Function: Afferent (sensory) axons relay changes in muscle length (distension) to the spinal cord. Mediate a variety of reflexes to maintain body posture and balance during movement. Also sends information used to gauge amount of effort required to perform certain movements.
- Muscle spindle:
-
- Tendon (Golgi) organs
- Location: Similar to muscle spindles but in the tendons near the insertion site with the muscle.
- Structure: Connective tissue components of muscle tissue contain both collagen fibers and elastic fibers and are continuous with the connective tissue of the tendons.
- Function: Afferent (sensory) axons relay stretching activity at the muscle-tendon(aka myotendinous) junction to the spinal cord to prevent damage from excessive tensile forces. Also sends information used to gauge amount of effort required to perform certain movements.
- Tendon (Golgi) organs
- Types of muscle fibers
- Type I aka slow aka red fibers; energy from oxidative phosphorylation (mitochondria); have lots of myoglobin for O2; prolonged continuous contraction;
- Type IIa aka fast intermediate; energy from a mix of ox-phos and glycogen (anaerobic glycolysis); rapid contractions and short bursts;
- Type IIb aka fast fibers aka white fibers; few mitochondria, but lots of glycogen, and rely on glycolysis; rapid contractions; fatigue quickly; e.g. eye muscles.
CARDIAC MUSCLE (Session Objectives #1, #6, #7)
- Cardiac muscles are comprised of cells that each have a single nucleus called cardiac myocytes.
- The individual cardiac myocytes are stuck to one another at their ends by intercalated discs. These discs provide a means to transmit contractile forces along the fiber and so have lots of adherens junctions and desmosomes.
- The discs also have gap junctions to allow the myocytes to be electrically coupled to coordinate their contractility.
- The structure and function of the actin and myosin fibrils is similar in cardiac muscle as it is in skeletal muscle. The sarcoplasmic reticulum and T-tubules are similar in cardiac myocytes as skeletal muscle fibers, except they form diads instead of triads.
- Cardiac myocytes contract spontaneously; the gap junction coupling among fibers leads to waves of coordinated contraction.
- Cardiac myocytes have many more mitochondria than skeletal muscle fibers.
- Cardiac myocytes are not all lined up like skeletal muscle fibers, but instead form a kind of branched network.
- The atrial cardiac myocytes also act as an endocrine organ and release atrial natriuretic hormone (ANH), and so these cells have “dense-core” vesicles.
- Like skeletal muscle, cardiac muscle cells are post-mitotic. Unlike skeletal muscle, cardiac muscle lacks a population of satellite cells and thus cannot regenerate following injury.
- Specialized cardiac muscle fibers, Purkinje fibers, conduct nerve impulses through the cardiac muscle to coordinate the cardiac muscle contraction.
SMOOTH MUSCLE (Session Objectives #1, #5, #6, #7)
- Smooth muscle exists as collections of individual spindle shaped mononucleated cells present primarily in the walls of blood vessels and hollow viscera.
- The actin and myosin fibers within the cytoplasm of smooth muscle cells are not as well organized as in either skeletal or cardiac muscle, so striations are not visible.
- The actin fibers attach to dense bodies (analogous to Z-bands) distributed through the cytoplasm or attached to the sarcolemma.
- T-tubules are absent in smooth muscle cells, but plasmalemma invaginations, called caveolae, may be functionally similar.
- The myosin filaments form cross-links with the actin filaments and increases in intracellular Ca+2 cause contraction, as in other types of muscle fibers; however, smooth muscle actin filaments lack troponin and instead the Ca+2 ions bind to a protein called calmodulin which activates myosin light chain kinase to initiate contraction via phosphorylation of the myosin.
- Smooth muscle fibers contain a specific type of actin, called smooth muscle actin, and they also have additional intermediate filaments, desmin.
- Smooth muscle cells are innervated primarily by the autonomic nervous system. Most blood vessels receive sympathetic innervation and respond to epinephrine. Smooth muscles also have acetylcholine receptors, but these are muscarinic rather than nicotinic receptors.
- Smooth muscle cells are often extensively coupled by gap junctions to pass electrical stimulation from cell to cell.
- Smooth muscle transduce force by attachments to surrounding extracellular matrix.
- Smooth muscle contraction is slower than other muscle types, but can be maintained for long periods of time.
HISTOLOGY ATLAS AND PRACTICE EXERCISES
Slide 1: Skeletal muscle in Longitudinal Section
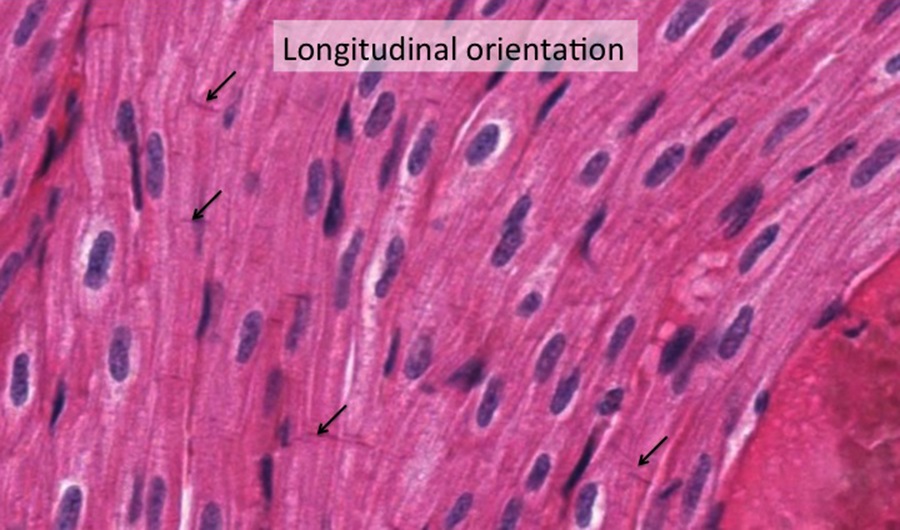
Skeletal muscle fibers form large solitary strips, display cross-striations and the round or oval nuclei are at the periphery of the fiber.

Above is an example of skeletal muscle cut in longitudinal section (along the long axis). At low magnification you can see the overall organization of a muscle; it looks fairly homogeneous but there are a few spaces between the fascicles of fibers that contain fat cells (these appear as white spaces, partly due to fixation artifact. The connective tissue encompassing the entire muscle is called the epimysium (highlighted in black), while the connective tissue surrounding groups of muscle fibers is called perimysium (highlighted in blue. The connective tissue surrounding individual muscle fibers within the fascicle is called endomysium. Also note the relative absence of branching of the fibers.
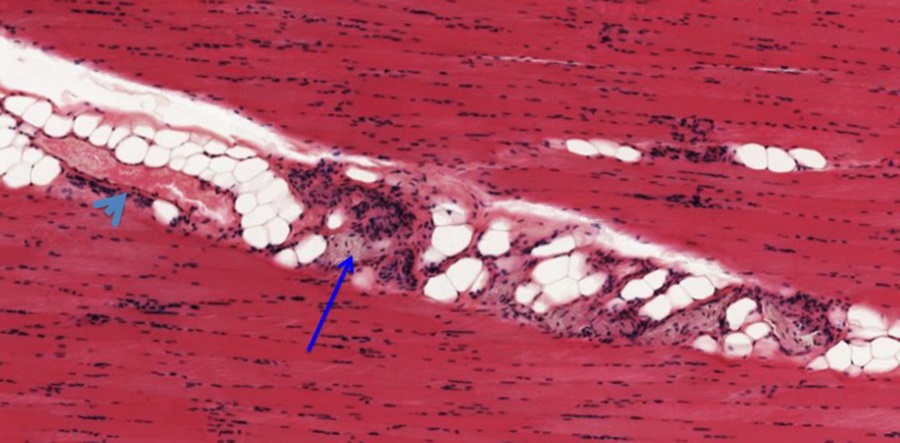
Nerves (blue arrow) and blood vessels (light blue arrowhead) are found in the perimysium layers seen at higher magnification above.

The micrograph above shows portions of muscle fibers cut longitudinally. The elongated nuclei are located in the periphery of the muscle fibers and comingle with nuclei of endothelial cells of capillaries that run between the fibers. Striations are evident within the muscle fibers. You should know the basis for the banding pattern observed in the striated muscle fibers and the organization of a sarcomere.
Optional activity: You can practice looking at a longitudinal sections of skeletal muscle using a virtual microscope with guiding text in the right panel at this link.
Slide 2: Skeletal Muscle in Cross-Section
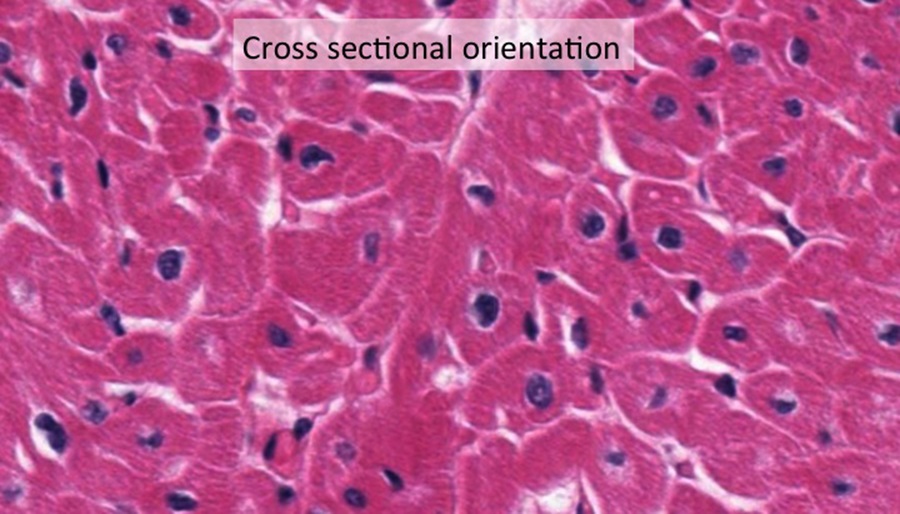
In cross-section of a skeletal muscle sample below, except for being larger than normal becuase of artifact shrinkage (white gaps between fibers), you can see where the endomysium would surround each individual muscle fiber.

At this high magnification the peripheral location of basophilic nuclei within the fiber is evident. Note the many capillaries (in cross section appear as small red circles adjacent to the muscle fibers; black arrows). The grainy appearance of the muscle fibers reflect myofibrils cut in cross section.
Optional activity: You can practice looking at a cross-sections of skeletal muscle using a virtual microscope with guiding text in the right panel at this link.
Slide 3: Cardiac Muscle

Above is a section through the heart; most of the cells are cardiac muscle fibers cut in various orientations, but some connective tissue (more lightly stained) can also be observed (arrows). For today, focus on the cardiac muscle cells and their characteristic features; we will see more of the heart and its structure later in the curriculum.
At higher magnification, cardiac muscle fibers can be distinguished from skeletal muscle fibers because they exist as single cells and do not fuse together to form muscle fibers like striated muscle cells; each cardiac muscle cell retains a centrally-located nucleus in the cell. Cardiac muscle cells are joined end-to-end with adjacent cardiac muscle cells by intercalated discs. These appear as delicate lines perpendicular to the long axis of cardiac muscle cells (Black arrows on image above). Gap junctions are one type of cell junction that can be found in the intercalated discs providing ionic continuity between cardiomyocytes. This continuity allows neighboring cariomyocytes to work together simultaneously as a coordinated syncytium. Cardiac muscle fibers are also branched, but this is sometimes harder to see. Cardiac muscle fibers also have striations, noticeable vaguely above, but they are not as distinct as seen in skeletal muscle fibers.
Optional activity: You can practice looking at a longitudinal sections of cardiac muscle using a virtual microscope with guiding text in the right panel at this link.
Compare the shape and position of the nuclei of the cross-sections of cardiomyocytes above with the picture of skeletal muscle in cross-section earlier in this chapter.
Optional activity: You can practice looking at a cross-sections of cardiac muscle using a virtual microscope with guiding text in the right panel at this link.
Slide 4: Smooth Muscle in the Ileum

The section above is through the ileum. The outer layers of the ileum are made of two layers of smooth muscle cells (indicated by the yellow band).
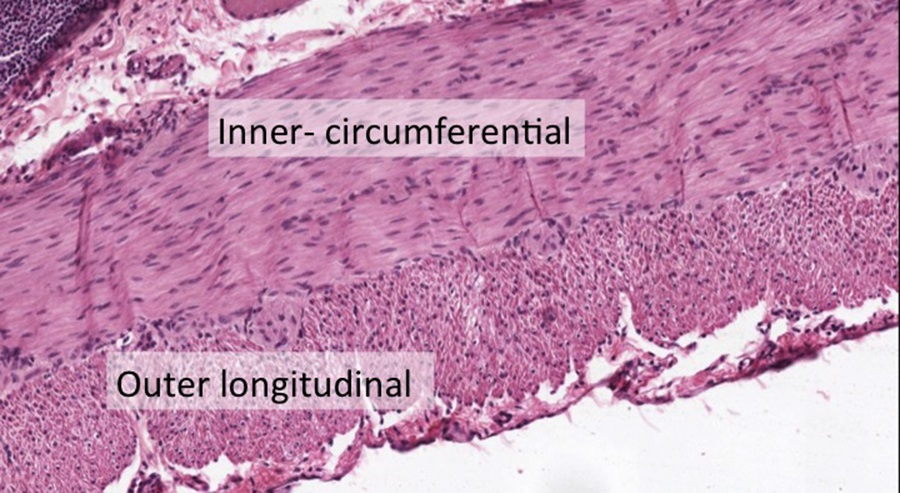
The inner layer of smooth muscle is circumferential around the ileum, while the outer layer runs along the long axis, ie. longitudinal, of the ileum. Because the section above is cut in cross-section (perpendicular to the length of the ileum), you are looking at what appears to be the opposite pattern as described. This section is therefore a convenient way to see smooth muscle cells cut in the two different orientations. Focus on the inner-circumferential layer when viewing the image below:
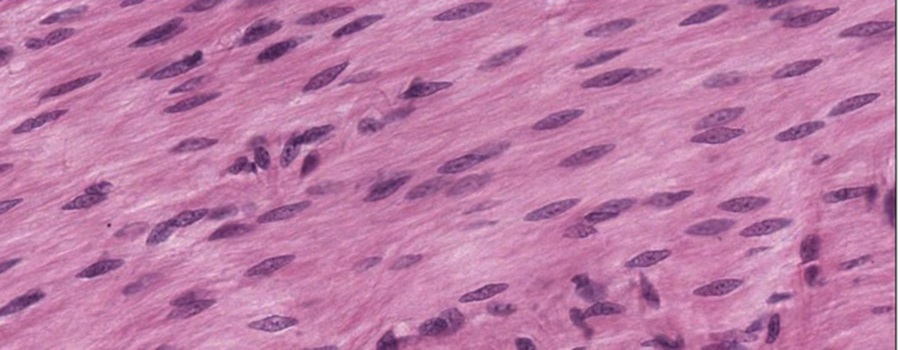
At higher magnification, you can see that the smooth muscle cells are small and spindly, they lack striations, and have slender, elongated nuclei when not in contraction that lie in the center of the cell (micrograph above). You would not mistake this for dense connective tissue because there are too many nuclei, indicating a cell-rich tissue (Session Objective #5). Smooth muscle cells lack the intercalated disks of cardiac cells or the striations of cardiac and skeletal muscle cells.
Optional activity: You can practice looking at a longitudinal sections of smooth muscle using a virtual microscope with guiding text in the right panel at this link. Notice that contraction of the smooth muscle cell in longitudinal section compresses the nucleus, giving it a cork-screw, or squiggly, shape.

In cross-section (above), smooth muscle fibers of the outer smooth muscle layer (longitudinal layer) are small and tightly packed circular profiles and occasionally contain centrally located nuclei.
Optional activity: You can practice looking at a cross-sections of smooth muscle using a virtual microscope with guiding text in the right panel at this link.
Slide 5: Smooth Muscle in the Oviduct
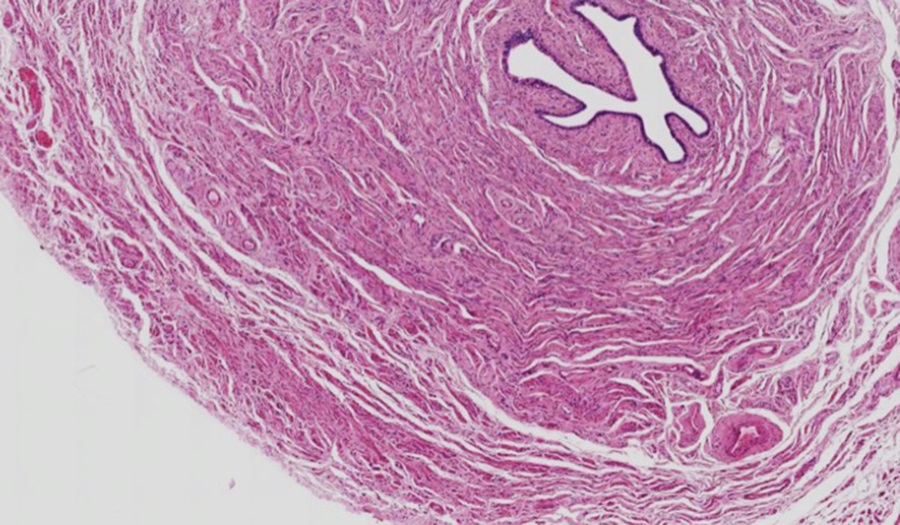
Above is a section through the oviduct, a thick smooth muscle tube lined by an epithelium. Examine the smooth muscle in longitudinal orientation (outer layer of the oviduct) in the micrograph below:

And in cross-section (inner layer of the oviduct) below:
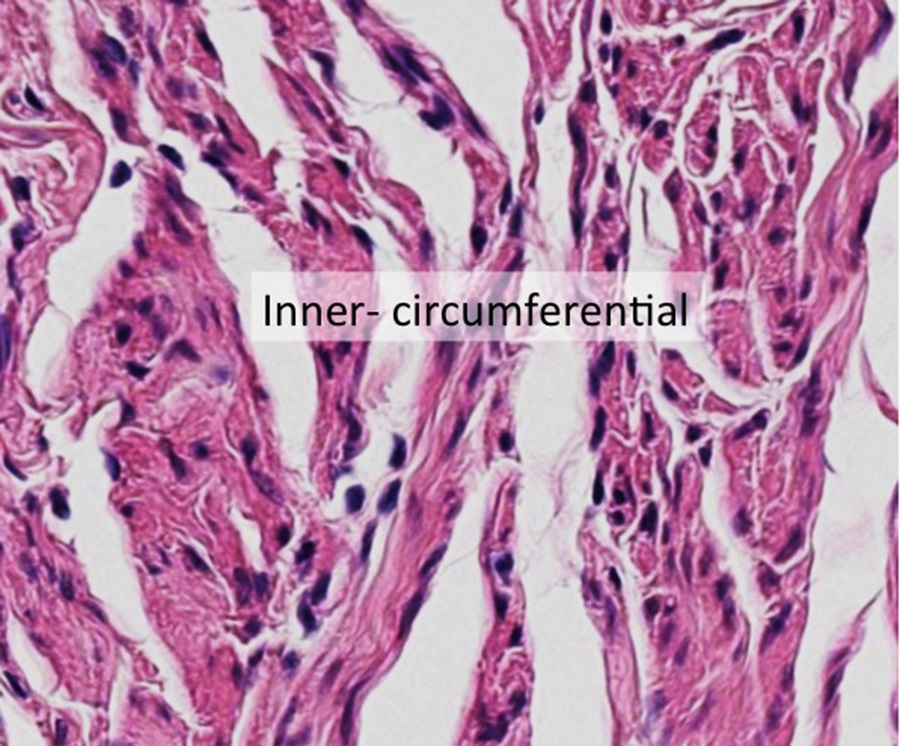
The smooth muscle cells are tapered at each end, but this is usually hard to see unless the cells are isolated.
Scanning through the low magnification micrograph of the oviduct section above (Slide 5), you may have spotted the artery in the lower right corner of the image. At higher magnification, the lumen of the blood vessel (asterisk, micrograph below) is filled with erythrocytes and is lined by simple squamous endothelium. Surrounding the epithelium is a layer of smooth muscle (SM), with a peripheral layer of connective tissue (CT). Note the difference in cellularity and the paucity of ground substance between smooth muscle and connective tissue below (session objective #5).
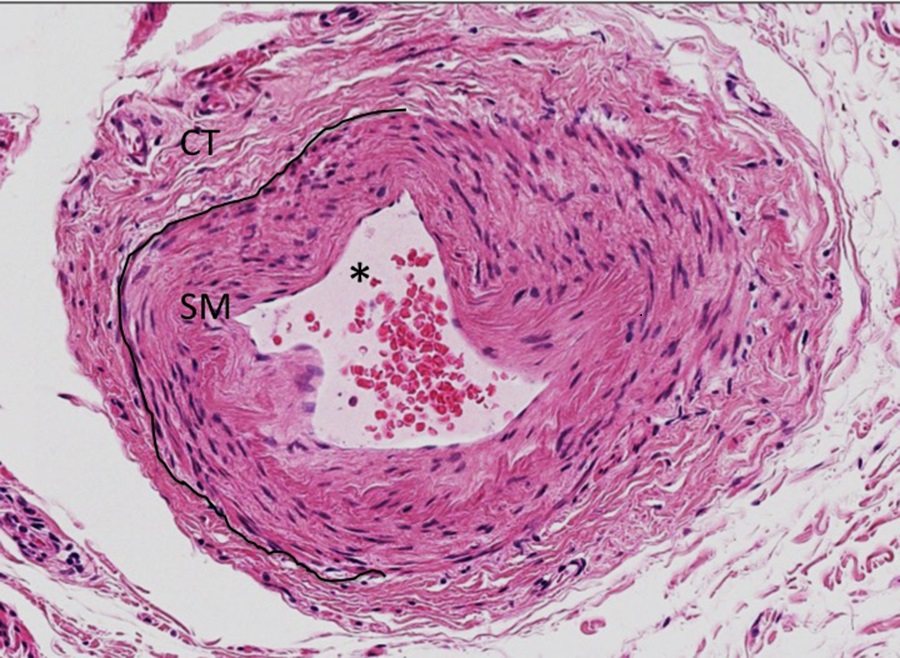
Slide 6: Smooth Muscle in the Esophagus
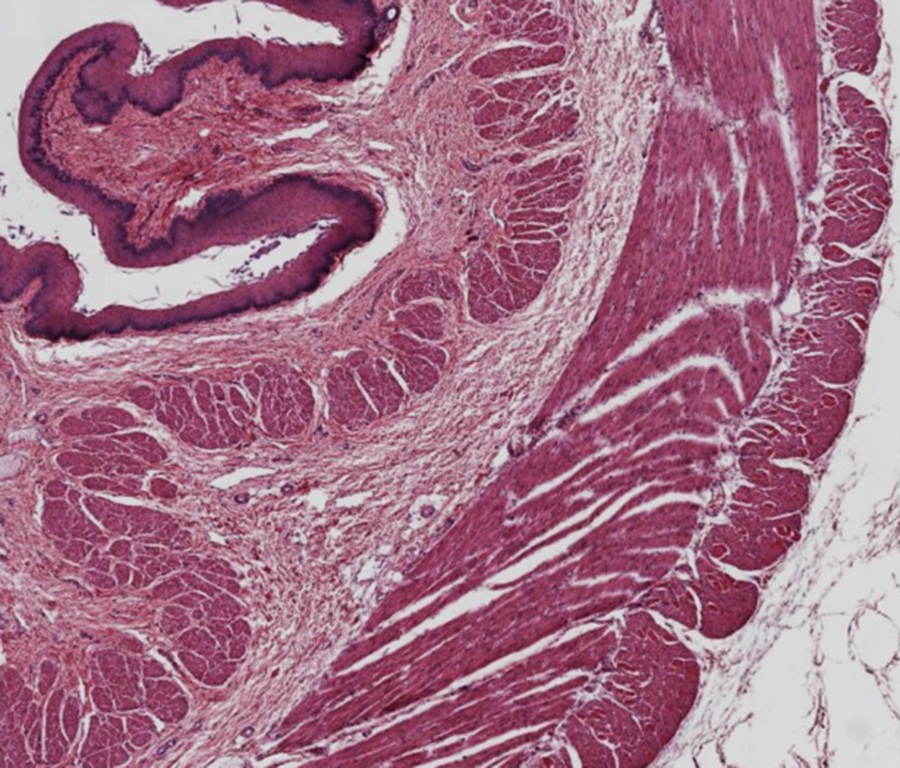
The muscular tube of the esophagus provides a way to compare the morphology of smooth and skeletal muscle cells. As the esophagus proceeds from the oral cavity to the stomach, the muscular composition changes from striated muscle near the oral cavity to a mixture of both, and finally to almost entirely smooth muscle as it approaches the stomach.
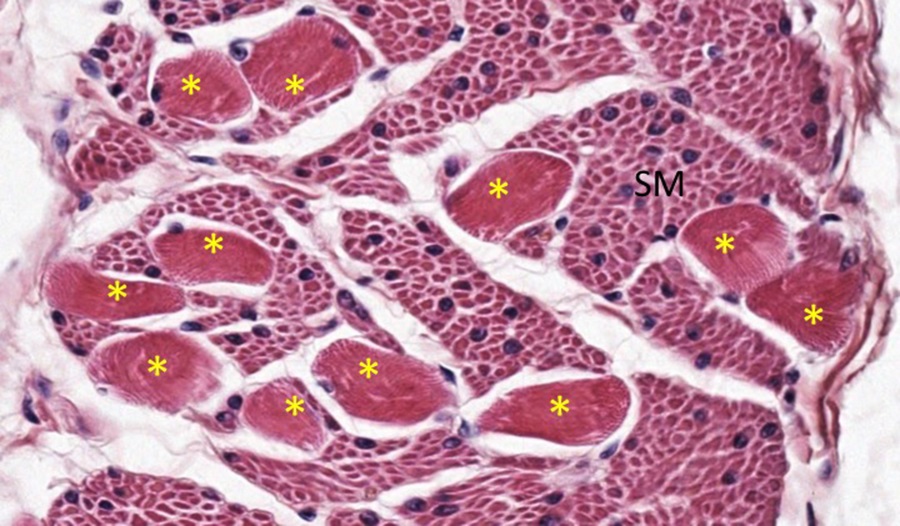
At higher magnification in the micrograph below, you can see a large amount of smooth muscle (SM) cells with a single round, more centrally-located nuclei compared to the scattered skeletal muscle fibers (asterisks) with multiple, eccentric nuclei and some evidence of granularity indicating sarcomeres (aka striations) in cross-section/oblique section.
The micrographs below have been incorrectly organized. Drag each micrograph to its correct row and column to compare the characteristic histological features of the three muscle types.
Drag Each Descriptive Text-box to Its Correct Muscle Column to Compare and Contrast Muscle Mechanics and Energy Use:
This Chapter’s PDF
- Note: The interactive features of this chapter are not reproducible in this PDF format.
ARCHIVAL PRE-CLASS MATERIALS FOR THIS SESSION:
- Pre-recorded videos: Muscle Histology
Feedback/Errata